
")

 |
 |
|||
| personajes | ||||
|
Alfonso Nápoles Gándara
|
|
|||
|
Alberto Barajas
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
En 1930, el misterioso azar nos reunió a un joven profesor de matemáticas y a mí en un palacio. Ese año sería crucial en nuestras vidas, pero creo que ninguno de los dos lo presentía, aun cuando yo lo escuchaba atentamente disertar sobre las variaciones del trinomio de segundo grado.
La Universidad estaba estrenando la autonomía que conquistó en la lucha de 1929. En los mexicanos duraba todavía el estremecimiento provocado por las elecciones de 1928, en las que un general menospreció la Constitución y quiso reelegirse matando a sus dos contrincantes para asegurar su triunfo, aunque a su vez, después fue víctima, de una mano fanática instrumento de la justicia inmanente. El general Obregón murió exactamente el 17 de julio de 1928, día de mi cumpleaños.
Entré a la preparatoria lleno de expectativas, pues esperaba respuesta a muchas preguntas que me inquietaban: ¿en qué clase de país me había tocado nacer?, ¿qué sentido tiene la vida humana?, ¿qué cosa es la universidad y el amor y la cultura? Me obsesionaba, sobre todo, la pregunta que se han hecho los adolescentes de todos los tiempos: ¿qué rumbo le daré a mi vida?
Cada día estaba más desorientado y no podía dormir. Por otro lado juzgaba a mis profesores con frialdad implacable ya que me parecían menos interesantes que algunos, magníficos, que había tenido en la Secundaria 3. Pero existían dos excepciones: el joven Alfonso Nápoles Gándara y el viejo Erasmo Castellanos Quinto. Sí, con ellos las clases de matemáticas y de literatura universal eran muy gratas.
Como Kafka, fatigué desesperado los corredores del Palacio de San Ildefonso, y súbitamente empecé a escuchar voces que parecían venir de algún confín remoto para calmar mis dolores metafísicos. Una voz me dijo al oído: “No existen sino anhelos, Barajas. Lo demás no existe, Por los menos no existe vitalmente. La realidad de que habla la ciencia es realidad pensada; realidad viva sólo la tienen los objetos cuando en ellos se prende nuestro deseo o nuestra nostalgia. Tener las cosas no nos importa, importa aspirar a ellas o extrañarlas cuando ya se han ido. Parecemos los hombres una caravana que camina bajo el sol insoportable del desierto. Nos hacemos la ilusión de que somos mercaderes, pero lo que en verdad queremos es sentir sed. Sed de saber, sed de amar. Sed de gozar de sufrir; de vivir... de morir...”
Otra me dijo algo semejante con una metáfora: “Quiero doblar el arco de la vida hasta que forme un círculo. De mis manos saldrá entonces la flecha de la existencia que persigo. Correrá entre los bosques, saltará sobre un río... El aire, herido por su vuelo, irradiará: con luz de constelaciones que viene del infinito. ¿Podrá el arco doblarse, sin romperse, hasta formar un círculo? La flecha que saldrá de mi arco ¿llegará a su destino?” El mundo empezaba a aclarárseme.
El profesor Nápoles caminaba muy aprisa y con la mirada hacia adelante. Siempre iba muy derecho pues tenía una de esas espaldas sin curvatura que sólo se ven en el Colegio Militar. Fuera de clase parecía no reconocernos, en cambio, en clase... tampoco. Sus exposiciones eran muy claras, ya que calculaba muy bien las dosis de conocimiento que podíamos absorber sin mayor esfuerzo. Escribía en el pizarrón lo que era necesario y suficiente, y no nos abrumaba con dictados inútiles. Su voz era muy clara y se escuchaba perfectamente hasta la última fila.
Un día empezamos a notarlo nervioso y preocupado, entonces nos anunció que el curso tendría que acortarse y el examen final sería en agosto. Por los periódicos supimos la causa de su inquietud: le acababan de conceder la beca Guggenheim para hacer estudios superiores en el Instituto Tecnológico de Massachusetts. Era el primer matemático mexicano que obtenía tal honor. A partir de ese momento, nuestro respeto por él dio un salto cuántico.
Al darnos las calificaciones del examen comentó brevemente mi trabajo. Su demostración es interesante, me dijo. Tiene diez. Esa vez fue el único que puso.
Para Nápoles fue seguramente muy angustioso el contacto brusco con el mundo de la ciencia internacional. En aquella época había una gran distancia entre la preparación matemática que se podía obtener en México y la que ofrecían las grandes universidades extranjeras. Sólo el que ha estado en ellas puede apreciar el esfuerzo heroico de Alfonso Nápoles para acreditar catorce cursos semestrales de matemáticas superiores de categoría A con la máxima calificación de H, aprobado con honor, en once de ellos. Si consideramos que esto ocurrió en un lapso de año y medio, su actuación fue muy notable.
Regresó a México en 1932 con un tesoro de conocimientos que ha compartido con sus discípulos y sin los cuales no hubiera sido posible la colaboración con grandes científicos extranjeros. Cuando yo discutía con Einstein una teoría rival a la suya, la de Birkhoff, pensaba que esa conversación hubiera sido imposible sin el curso de Cálculo Tensorial que tomé con don Alfonso. Al pensar en su vida recuerdo inevitablemente las palabras que también escuché en 1930, mi año terrible, y que ya he mencionado en alguna otra ocasión: “Hay hombres que sólo entran al combate cuando el Rey está mirando. O son como Aristo, aquel filósofo tan elegante que sólo filosofaba cuando sus amigos lo llevaban en una litera lujosa. Hay otros, en cambio, que trabajan siempre que se necesita y en las condiciones menos favorables; dispuestos todo el tiempo a cumplir con sus deberes y sus ideales”.
La biografía de Nápoles nos muestra su devoción por las matemáticas y por la Universidad. Esta prodigiosa Universidad de México que tanto me deconcertó al principio y que al fin me reveló cuál era mi destino, mi dharma. Si en un momento de crisis no hubiera tenido un profesor de matemáticas tan distinguido, probablemente habría seguido otra carrera.
Alfonso Nápoles Gándara nació en Cuernavaca el 14 de octubre de 1897. Allí estudio la primaria; después estuvo en la Escuela Nacional Preparatoria y en la Escuela Nacional de Ingenieros, donde por su talento y originalidad llamó la atención del extraordinario Sotero Prieto.
Cuando Sotero murió, asumío el liderazgo del movimiento matemático en México. Más adelante, formó parte del grupo fundador de la actual Facultad de Ciencias, donde dirigió el Departamento de Matemáticas de 1939 a 1965. En 1942 se fundó el Instituto de Matemáticas, del que fue su primer director en noviembre de 1942, inició en Saltillo la serie de congresos de matemáticas que se han venido realizando con tanto éxito a lo largo de muchos años. Como consecuencia del primer congreso se creó la Sociedad Matemática Mexicana, la cual presidió hasta 1961. Desde entonces es Presidente Honorario Vitalicio.
Está casado con la señora Guadalupe Salazar y su hijo es el arquitecto Alfonso Nápoles Salazar, es hombre muy inteligente y muy creativo, quien además es distinguido y respetado en su profesión.
En fin, como quería el poeta, don Alfonso supo empuñar el arco con valentía y firmeza para lograr que la flecha llegara a su destino.
Septiembre de 1991
|
||||
|
Alberto Barajas
Instituto de Matemáticas,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Barajas Celis, Alberto.(1999). Alfonso Nápoles Gándara. Ciencias 53, enero-marzo, 28-30. [En línea]
|
||||
|
|
||||
|
El diablo de los números
|
 |
|
|
Hans Magnus Enzensberger
Ediciones Siruela, España, 1998
|
||
| HTML ↓ | ← Regresar al índice / artículo siguiente → | |
|
Hacía mucho que Robert estaba harto de soñar. Se decía: Siempre me toca hacer el papel de tonto.
Por ejemplo, en sueños le ocurría a menudo ser tragado por un pez gigantesco y desagradable, y cuando esta a punto de ocurrir llegaba a su nariz un olor terrible. O se deslizaba cada vez más hondo por un interminable tobogán. Ya podía gritar cuanto quisiera ¡Alto! o ¡Socorro!, bajaba más y más rápido, hasta despertar bañado en sudor.
A Robert le jugaban otra mala pasada cuando ansiaba mucho algo, por ejemplo una bici de carreras con por lo menos veintiocho marchas. Entonces soñaba que la bici, pintada en color lila metálico, estaba esperándole en el sótano. Era un sueño de increíble exactitud. Ahí estaba la bici, a la izquierda del botellero, y él sabía incluso la combinación del candado: 12345. ¡Recordarla era un juego de niños! En mitad de la noche Robert se despertaba, cogía medio dormido la llave de su estante, bajaba, en pijama y tambaleándose, los cuatro escalones y... ¿qué encontraba a la izquierda del botellero? Un ratón muerto. ¡Era una estafa! Un truco de lo más miserable.
Con el tiempo, Robert descubrió cómo defenderse de tales maldades. En cuanto le venía un mal sueño pensaba a toda prisa, sin despertar. Ahí está otra vez este viejo y nauseabundo pescado. Sé muy bien qué va a pasar ahora. Quiere engullirme. Pero está clarísimo que se trata de un pez soñado que, naturalmente, sólo puede tragarme en sueños, nada más. O pensaba: Ya vuelvo a escurrirme por el tobogán, no hay nada que hacer, no puedo parar de ningún modo, pero no estoy bajando de verdad.
Y en cuanto aparecía de nuevo la maravillosa bici de carreras, o un juego para ordenador que quería tener a toda costa —ahí estaba, bien visible, a su alcance, al lado del teléfono—, Robert sabía que otra vez era puro engaño. No volvió a prestar atención a la bici. Simplemente la dejaba allí. Pero, por mucha astucia que le echara, todo aquello seguía siendo bastante molesto, y por eso no había quien le hablara de sus sueños.
Hasta que un día apareció el diablo de los números.
Robert se alegró de no soñar esta vez con un pez hambriento, y de no deslizarse por un interminable tobogán desde una torre muy alta y muy vacilante. En su lugar, soñó con un pradera. Lo curioso es que la hierba era altísima, tan alta que a Robert le llegaba al hombro y a veces hasta la cabeza. Miró a su alrededor y vio, justo delante de él, a un señor bastante viejo, bastante bajito, más o menos como un saltamontes, que se mecía sobre la hoja de acedera y le miraba con ojos brillantes.
—¿Quién eres tú?— preguntó Robert.
El hombre le gritó, sorprendentemente alto:
—¡Soy el diablo de los números!
Pero Robert no estaba de humor para aguantarle nada a semejante enano.
—En primer lugar —dijo—, no hay ningún diablo de los números.
—¿Ah, no? ¿Entonces por qué estás hablando conmigo, si ni siquiera existo?
—Y en segundo lugar, odio todo lo que tiene que ver con las Matemáticas.
—¿Por qué?
—“Si dos panaderos hacen 444 trenzas en seis horas, ¿cuánto tiempo necesitarán cinco panaderos para hacer 88 trenzas?” Qué idiotez —siguió despotricando Robert—. Una forma idiota de matar el tiempo. Así que ¡esfúmate! ¡Largo!
El diablo de los números se bajó con un elegante salto de su hoja de acedera y se sentó al lado de Robert, que en protesta se había sentado entre la hierba, alta como un árbol.
—¿De dónde te has sacado esa historia de las trenzas? Seguro que del colegio.
—¡Y de dónde si no! —dijo Robert—. El señor Bockel, ese principiante que nos da Matemáticas, siempre tiene hambre, a pesar de estar tan gordo. Cuando cree que no le vemos porque estamos haciendo los deberes, saca una trenza de su maletín y se la devora mientras nosotros hacemos cuentas.
—¡Vaya! —exclamó el diablo de los números, sonriendo con sorna—. No quiero decir nada en contra de tu profesor, pero la verdad es que eso no tiene nada que ver con las Matemáticas. ¿Sabes una cosa? La mayoría de los verdaderos matemáticos no sabe hacer cuentas. Además, les da pena perder el tiempo haciéndolas, para eso están las calculadoras. ¿No tienes una?
—Sí, pero en el colegio no nos dejan usarla.
—Ajá —dijo el diablo de los números—. No importa. No hay nada que objetar a un poco de práctica con las tablas. Puede ser muy útil si uno se queda sin pilas. ¡Pero las Matemáticas, ratoncito, eso es muy diferente!
_______________________________________________________________
como citar este artículo → Magnus Enzensberger, Hans. (1999). El diablo de los números. Ciencias 53, enero-marzo, 68. [En línea] |
||
|
|
← Regresar al índice / artículo siguiente → | |
 |
 |
|||||
|
El lirio acuático, ¿una planta nativa
de México?
|
|
|||||
|
María Guadalupe Miranda A. y Antonio Lot Helgueras
conoce más del autor
|
||||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||||
|
El lirio acuático (Eichhornia crassipes) es una planta libre flotadora miembro de la familia de las Pontederiaceae, que ocupa un lugar sobresaliente entre las comunidades de hidrófitas de agua dulce de las regiones tropicales y subtropicales del mundo. Las flores que la conforman, por su forma y color, son tanto la estructura más vistosa de la planta como el principal motivo que ha favorecido su introducción en estanques y acuarios de todo el mundo. Los lóbulos de la flor presentan tonalidades que van del color morado al lila y al azul claro, destacando en el lóbulo superior central una mancha de color amarillo.
Su distribución en México es sumamente amplia, ya que crece en una gran diversidad de hábitats dulceacuícolas localizados desde el nivel del mar hasta los 2 250 metros de altitud. Los únicos lugares en los que no se ha registrado son los estados de Baja California Sur, Chihuahua, Zacatecas, Tlaxcala y Yucatán.
La abundancia y la frecuencia con la que encontramos el lirio acuático (Eichhornia crassipes) en los cuerpos de agua naturales y artificiales en México nos hace pensar que se trata de una planta nativa del país.
En las regiones fuera de su área natural de distribución, el lirio acuático es considerado maleza (planta indeseable) por la serie de problemas que acarrea su presencia y, sobre todo, por el crecimiento tan rápido de su población. En hábitats perturbados sustituye con frecuencia a elementos de la flora nativa. Tiene diversos efectos perjudiciales, entre los que se pueden mencionar el entorpecimiento de la navegación por ríos y lagunas, la elevación de los índices de evaporación y el impedimento del paso de la luz al fondo de los estanques, lo que trae como consecuencia la eliminación de microalgas que son el alimento de crustáceos y peces. Sabemos que el lirio acuático es una de las malezas, o planta invasora, más difíciles de exterminar en áreas inundables manejadas por el hombre en las regiones tropicales y subtropicales del mundo.
Según Orozco-Segovia y Vázquez-Yanes esta planta es considerada nativa de México y de Centroamérica, aunque existen dudas sobre las causas de su distribución fuera de las cuencas del Amazonas y del Orinoco, que corresponden al centro de origen y de distribución primaria, aunque estos autores también señalan que su presencia en México podría deberse a una introducción temprana.
En la búsqueda que realizamos en diversas fuentes botánicas antiguas del país hallamos algunos datos que parecen apuntar hacia la segunda suposición. En la Historia de las cosas de la Nueva España, obra escrita por Fray Bernardino de Sahagún, publicada cerca de 1590, se hace referencia a las plantas que existían en los lagos, pero no se incluye al lirio acuático. Igualmente, en la Historia natural de la Nueva España, publicada en 1576 por el naturalista Francisco Hernández, protomédico del rey Felipe II, se mencionan algunas plantas acuáticas, pero, una vez más, no al lirio acuático. En el Códice Badiano, escrito en 1552 reconocimos íconos de macrofitas acuáticas, pero el lirio acuático tampoco aparece.
Buscando las plantas útiles del México antiguo, Torres revisó en 1985 otras obras del siglo XVI, como las crónicas de José Acosta, Antonio de Ciudad Real, Fray Diego de Landa y Fray Alonso de Molina, en donde no se hace ninguna referencia al lirio acuático.
Algunos estudios palinológicos recientes que se han realizado en la zona central de México con objeto de encontrar registros de polen de lirio acuático en estratos correspondientes a épocas precolombinas no han tenido éxito. Todo esto nos hace pensar que el lirio acuático no formaba parte de la flora acuática de México en esa época.
En 1882, en el Calendario botánico del valle de México, Bárcena tampoco menciona al lirio acuático. El primer registro lo encontramos hasta 1903, en la obra de Manuel Urbina, después de la cual otros autores como Reiche, Ramírez-Cantú y Conzatti ya lo consideran parte de la flora acuática mexicana.
El origen
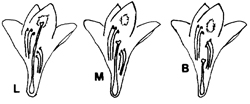
En el lirio acuático la morfología del órgano femenino de la flor o estilo varía en longitud y recibe los nombres de longistila (L), mesostila (M) y brevistila (B), respectivamente de mayor a menor talla (figura 1). A este fenómeno se le conoce como “heterostilia trimorfa” o “tristilia”.
Asumiendo que la mayor diversidad de formas de las especies se encuentra en la cercanía al centro de origen y de dispersión, y sin descartar la influencia del ambiente en la manifestación de la característica, podemos concluir que el lirio acuático es originario de la cuenca del Amazonas, en Brasil, ya que es únicamente allí donde se ha reportado la “tristilia”. También hay registros de “tristilia” en lagos cercanos a la confluencia de los ríos Paraguay y Paraná, en Argentina, en donde posiblemente se dispersó por medios naturales.
Las poblaciones de lirio acuático en Guyana, Venezuela y Colombia, así como en el Caribe, América Central y zonas templadas de América del Norte, parecen ser resultado de introducciones, ya que carecen del tamaño intermedio (la forma B). Esta idea se refuerza por la carencia de polinizadores especializados en América Central. El polinizador especializado registrado para el lirio es una abeja con lengua larga (Ancyloscelis gigas), que se ubica únicamente en la región amazónica donde se presenta la “tristilia”.
Resulta pertinente preguntarse si el lirio acuático pudo haber llegado a Centroamérica y a Norteamérica a través de mecanismos de distribución natural. Los órganos vegetativos de dispersión del lirio acuático incluyen las plantas completas u otras estructuras, como rizomas y estolones, y sólo pueden colonizar nuevos sitios que estén inundados. La dispersión es resultado del movimiento de los órganos vegetativos en los ríos, o bien, de las inundaciones que pueden acarrear los propágulos a lagos adyacentes, canales o diques.
Otro mecanismo de transporte de las semillas de algunas plantas acuáticas es el viento, pero las distancias alcanzadas son muy cortas. En el caso del lirio acuático, es poco probable que el viento las disperse, pues no tienen estructuras de transporte aéreo, como sucede con el tule ancho (Typha) y el carrizo (Phragmites).
Desde hace un siglo, Darwin explicó que la amplia distribución de ciertas especies vegetales de agua dulce se debía al acarreo de semillas y fragmentos vegetativos sobre el plumaje y las patas de las aves acuáticas. No obstante, para el caso del lirio acuático, que como ya mencionamos es una planta originaria de Brasil, es riesgoso considerar que este mecanismo como sea el responsable de las poblaciones del lirio asentados en Centro y Norteamérica, pues se carece de pruebas que indiquen su presencia desde el pasado.
Sin embargo, el hombre, de manera intencional, ha intervenido en la introducción y en la naturalización de nuevas plantas entre los hemisferios y entre los continentes. Sobre el lirio acuático, gracias a la magnífica obra compiladora de Gopal, se han documentado numerosos casos de introducciones a través de jardines botánicos instalados a partir de diversas exploraciones de naturalistas europeos en la región amazónica, en donde lo atractivo de su inflorescencia llamó considerablemente la atención.
Uno de los casos de introducción y naturalización del lirio acuático mejor conocidos sucedió en Norteamérica en 1884, con motivo de una exhibición de horticultura en la exposición de algodón en Nueva Orleans. Posteriormente, hacia 1894, en el delta del río Mississipi en Lousiana se registró el primer obstáculo serio para navegar en los ríos y corrientes que desembocaban en el golfo de México. Se considera que la naturalización del lirio acuático en esa región de Estados Unidos ocurrió durante la década de 1860 a 1870, y que hacia 1880 y, claramente después de 1900, el lirio acuático ya había invadido otras regiones como los Everglades en Florida. Más adelante, en 1903, el lirio se colectó por primera vez en Texas y, al año siguiente, ya se encontraba en California.
En Las chinampas del Distrito Federal, de Santamaría, se hace referencia a la introducción del lirio acuático a México: “entre las plantas acuáticas destaca el lirio acuático que es conocido desde hace solo catorce años, en que fue introducido por orden del general Carlos Pacheco, entonces secretario de Fomento”. De acuerdo con esta información, fue en 1898 cuando el lirio acuático llegó a los lagos del valle de México.
También debe considerarse la dispersión accidental, en particular de las semillas y de las fracciones vegetativas que crecen entre los cultivos irrigados y que se distribuyen con los productos cosechados y a través de embarcaciones, vehículos y maquinaria agrícola.
Su fácil dispersión
La dispersión del lirio y su establecimiento exitoso están directamente relacionados con las características ambientales abióticas y bióticas. A pesar de la amplia tolerancia a los factores abióticos del medio, y de ser una especie que puede llegar a invadir cuerpos de agua en regiones templadas y a altitudes considerables (su registro a mayor altitud en el mundo es en la ciudad de México), el lirio acuático no soporta las heladas y su crecimiento es limitado por la salinidad. En todo el mundo, su crecimiento óptimo se da a pH de 7 y adquiere el rango de infestación masiva entre 6.2 y 7.
En la variedad de hábitats de los sistemas fluviales del Amazonas y de los ecosistemas acuáticos de la región neotropical, en general, el lirio compite con otras especies de hidrófitas libres flotadoras (Azolla spp., Hydromistria stolo nífera, Pistia stratiotes, Lemna spp., Salvinia spp.) en mosaicos densos típicos de este tipo de comunidades naturales. Es interesante notar que el crecimiento masivo y con características de infestación de cualquiera de las especies mencionadas (especialmente Salvinia) en sus áreas naturales de distribución, sólo se ha registrado en donde artificialmente se cambió el régimen hidrológico, favoreciendo la formación de poblaciones dominadas por una sola especie.
La química del agua no limita el crecimiento del lirio acuático, pues su rango de tolerancia es amplio y el único factor que lo afecta es la carencia de fósforo. El lirio puede establecerse en aguas con pocos nutrientes, debido a las asociaciones microbianas que presenta en la rizosfera, y mientras más eutrófico sea el medio, más éxito tiene su propagación.
Entre las características de adaptación más importantes del lirio acuático están: a) su rápida capacidad de reproducción vegetativa; b) la habilidad para regenerarse a partir de pequeñas porciones del talo vegetativo; c) la completa o parcial independencia de la reproducción sexual; d) la morfología del crecimiento, que resulta en el desarrollo de la mayor área posible de tejido fotosintético, con relación al total de la planta y el cual rápidamente ocupa la superficie del agua, y e) independencia de las condiciones del sustrato y del nivel del agua.
Existen especies taxonómicamente cercanas, las cuales no son invasoras. Así, Eichhornia azurea es una especie estrechamente cercana a E. crassipes, con patrones de distribución nativa similar. Además, en contraste al hábito libre flotador de E. crassipes, E. azurea y las otras seis especies del género están unidas al sustrato. Es claro que la habilidad para llevar a cabo una propagación vegetativa rápida, unida al alto grado de movilidad, ha favorecido la dispersión de E. crassipes por encima de E. azurea.
Las fluctuaciones del nivel del agua inducen al lirio acuático a adoptar una forma emergente, con la elongación del pecíolo. Cuando la desecación es larga, los estolones quedan entrelazados esperando la época de inundación. A esta capacidad para modificar la forma y el tamaño de sus estructuras vegetativas cuando crecen bajo diferentes condiciones ambientales se le denomina plasticidad morfológica. Este atributo se ejemplifica bien en el lirio con la “heterofilia” o con la presencia en un solo individuo de dos o más tipos diferentes de hojas. La plasticidad morfológica y la habilidad para experimentar un crecimiento vegetativo rápido son los atributos clave que le dan a Eichhornia crassipes el éxito en la competencia interespecífica con otras plantas acuáticas invasoras.
Como en otras muchas macrofitas, en el lirio acuático hay una tendencia a reemplazar la reproducción sexual por la asexual. El ciclo completo de semilla a semilla es muy lento, ya que dura más de cinco meses. Además, para la germinación de la semilla es un requisito indispensable, la escarificación (rompimiento de la cubierta externa). En la propagación vegetativa el lirio produce estolones (tallos horizontales) que desarrollarán hojas arrosetadas de una yema terminal. Este proceso se repite en las plantas hijas y cuando se tienen las condiciones ideales, un número inmenso de plantas se pueden producir en corto tiempo, hasta duplicar la población en un periodo que va de cinco a quince días.
La presencia masiva e invasora del lirio ha llevado a buscar soluciones para su control. El hecho de que no existan herbívoros naturales, ni plagas que lo afecten, ha llevado a que actualmente se realicen estudios para utilizar hongos patógenos que puedan limitarlo. También se estudian diferentes artrópodos (coleópteros y lepidópteros) que puedan eliminarlo y se han probado, además, peces herbívoros como control biológico, entre los que podemos mencionar: Ctenopharyngodon idella, Tilapia rendalli y Metynnis roosevelti.
Una planta introducida
A través de los registros históricos, antropológicos y palinológicos, no se ha podido comprobar la existencia del lirio acuático en México en la época precolombina, por lo que se podría pensar que fue introducido voluntariamente como planta de ornato en Estados Unidos hacia 1884. Es posible que de ahí se haya dispersado en forma natural por medio de aves migratorias hasta llegar a nuestro país.
De cualquier forma, su introducción o naturalización en los lagos mexicanos no debió ocurrir antes de 1884. Lo que si es claro es su establecimiento y creciente poblamiento en nuevos espacios, desplazando a otras plantas acuáticas más sensibles a la perturbación humana.
La presencia del lirio acuático en ecosistemas lacustres estables con una alta biodiversidad es un fenómeno raro y su condición de planta “oportunista” sólo ocurre con la alteración del hábitat, especialmente cuando se presenta un incremento de nutrientes (eutroficación) por el derrame de fertilizantes o aguas negras. Desafortunadamente, hoy día es alarmante la modificación de los ambientes acuáticos naturales y el aumento de cuerpos de agua artificiales (presas, canales de irrigación, estanques de crecimiento, etcétera) que, por su inevitable conexión, pueden eventualmente contaminar grandes sistemas acuáticos a través de los ríos y de las crecientes durante los meses de lluvias y con la presencia de ciclones y otras perturbaciones meteorológicas. Con ello se favorece la invasión de Eichhornia crassipes como maleza a niveles que no pueden de controlar ni la naturaleza ni el hombre.
|
||||||
|
Referencias Bibliográficas
Gopal, B. 1987. Water Hyacinth. Aquatic Plant Studies. 1. Amsterdam, Elsevier Science Publishers, 471 p.
Mitchell, D. 1974. Aquatic vegetation and its use and control. Nueva York, unesco. pp. 38-49.
Orozco-Segovia, A. y C. Vázquez-Yáñez. 1993. Especies Invasoras: su impacto sobre las comunidades bióticas. México, Pronatura, A.C. pp. 29-31.
Sculthorpe, C. 1967. The biology of aquatic vascular plants. Londres, Edward Arnold. 610 p.
West, R. y P. Armillas. 1959. “Las chinampas de México, poesía, y realidad de los jardines flotantes”, en Cuadernos Americanos, 50, pp. 165-182.
|
||||||
|
María Guadalupe Miranda A.
UAM, Iztapalapa.
Antonio Lot Helgueras
Instituto de Biología,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Miranda A., María Guadalupe y Lot Helgueras, Antonio. (1999). El lirio acuático, ¿una planta nativa de México? Ciencias 53, enero-marzo, 50-54. [En línea] |
||||||
|
|
||||||
El método de las ciencias
 |
||
|
Rosaura Ruiz y Francisco J. Ayala
Fondo de Cultura Económica, México, 1998.
|
||
| HTML ↓ | ← Regresar al índice / artículo siguiente → | |
|
La ciencia se mueve, cambia de lugar, se agita. Es veleidosa y frágil. Por dentro y por fuera, se transforma. Está tan viva como un ejemplar de una especie, libre o en cautiverio, que muestra facetas desconocidas aun para los mismos que, en apariencia, la elaboran con absoluto control, y se les escapa, huye. Y luego se queda quieta, inmóvil, camuflada, a la espera, como si supiera que una infinidad de miradas la acechan, pendientes de ella, para aprehenderla. Los científicos saben todo esto y saben también que la certeza absoluta no es inherente al método científico, no es infalible; avanza cuando unos científicos refutan teorías de otros científicos, o de sí mismos al reconocer equivocaciones, y entonces aquél adquiere uno de sus rasgos más significativos, el lado humano, en el momento en que hace que la ciencia avance, socializada por vía del lenguaje. He aquí el tema central de esta obra de Ruiz y Ayala: a la ciencia se la puede estudiar y comprender de igual modo que como han procedido las epistemologías evolucionistas, sólo que en este caso no son los entes vivos la unidad de conocimiento, sino los conceptos y las teorías, en su origen, devenir y futuro, partiendo de la idea de que todo términos darwinistas, aunque por supuesto, en la aproximación de este problema es necesario adecuar el darwinismo a la teorización de carácter social, ya que los primeros darwinistas también pensaron que su paradigma era tan general que podía extenderse a toda entidad cambiante. Así, las maneras discrepantes en que varias teorías tratan de dar cuenta de determinado problema —específicamente el desarrollo de la ciencia, con sus controversias y polémicas— se analizan en estas páginas en las que el propósito más importante de los autores es estimular el interés en estas teorías, reconocer sus fundamentos, identificar lo que hay de válido en ellas y definir sus límites.
_______________________________________________________________
como citar este artículo → Ruiz Gutiérrez, Rosaura y Ayala, Francisco J. (1999). El método de las ciencias. Ciencias 53, enero-marzo, 69. [En línea]
|
||
|
|
← Regresar al índice / artículo siguiente → | |
Energía
 |
||
|
Ana María Sánchez, María Trigueros y
Julia Tagüeña
Historias de la ciencia y la técnica,
UNAM, México, 1999.
|
||
| HTML ↓ | ← Regresar al índice | |
|
Energía... Cuántas veces hemos oído esta palabra. Aunque la usamos muy a menudo, ¿podríamos decir qué es?
La energía es un concepto complejo de definir pero muy útil. Lo utilizan casi todas las ciencias y es también parte de nuestro vocabulario cotidiano.
¿Hace cuánto tiempo que hablamos de energía? Estamos tan habituados a hacerlo que pensamos que el concepto es muy antiguo, pero no es así. Los antiguos griegos usaban la palabra “energía” para referirse a la eficacia, el poder o la virtud para obrar. Ni Galileo ni Newton, siglos después, conocían el concepto en la forma en que hoy lo manejamos. No fue sino hasta mediados del siglo xix cuando varios científicos que hacían experimentos diferentes en diversos lugares encontraron que fenómenos que hasta entonces se pensaban ajenos unos a otros, como el calor y el movimiento, la electricidad y el magnetismo, el movimiento y la luz, la afinidad química y el calor, y otros más, se relacionaban entre sí. Es más, estos fenómenos podían transformarse uno en el otro y en esa transformación podía definirse un concepto abstracto que da cuenta de algo que se conserva: la energía.
La energía es un concepto unificador en la naturaleza, y por su carácter abstracto es más fácil de definir en términos de sus manifestaciones.
La energía se manifiesta de muchas formas que pueden intercambiarse: mecánica, calorífica, eléctrica, química, magnética, nuclear. Las distintas formas de energía se transforman unas en otras, y lo más importante es que en ese proceso la cantidad total de energía se conserva. A este fenómeno se le llama principio de conservación de la energía.
El descubrimiento del principio de conservación de la energía representó un momento importante en la historia de las ciencias. Su nacimiento permitió descubrir que varias de ellas, que hasta ese momento se estudiaban por separado, constituían partes de un todo: la física. Desde su nacimiento este principio ha estado ligado íntimamente a la tecnología. Su aplicación permitió el desarrollo de nuevas industrias que cambiaron totalmente nuestra forma de vida.
El concepto de energía ha sido un motor del desarrollo de la ciencia y la tecnología. Su papel ha sido fundamental y sólo mediante un conocimiento profundo de este concepto y de sus consecuencias podremos comprender y manejar los efectos de la tecnología sobre el ambiente y la sociedad.
_______________________________________________________________
como citar este artículo → Sánchez, Ana María y Trigueros María, Tagüeña Julia. (1999). Energía. Ciencias 53, enero-marzo, 70. [En línea]
|
||
|
|
← Regresar al índice | |
 |
 |
|||
| del bestiario |
||||
| La biodiversidad en el Asteroide B 612 |
|
|||
|
Héctor T. Arita
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
Si existiera un programa de conservación de la biodiversidad a nivel del sistema solar, el asteroide B 612 difícilmente se contaría entre las áreas prioritarias para establecer una reserva biológica. Efectivamente, si un hipotético exobiólogo realizara el inventario del planetoide hogar del Principito de Saint-Exupery, se encontraría con una flora representada por una vanidosa flor encerrada en un domo protector, unas cuantas flores de otra especie no muy bien determinada y unos cuantos brotes de baobab. Asimismo, encontraría que no existe fauna nativa en el lugar, y que el único animal es un borreguito encerrado en una caja. El asteroide no entraría, definitivamente, en las listas de sitios de megadiversidad.
El término “megadiversidad” fue acuñado hace algunos años por el biólogo estadounidense Russell Mittermeier para definir a la docena de países en los que se concentra la mayor parte de la diversidad biológica del planeta. Para identificar estos países megadiversos, Mittermeier recopiló información sobre el número de especies de los principales grupos biológicos y constató que existen unas pocas naciones que sobresalen en casi todos los rubros. México, por ejemplo, está incluido en este selecto grupo de países porque tanto su flora de plantas vasculares como sus faunas de reptiles, anfibios y mamíferos se encuentran entre las diez más ricas del planeta en número de especies. La riqueza de especies, es decir, el total de especies de plantas o animales que se encuentran en un sitio dado, es un criterio relativamente fácil de medir e interpretar, y en muchos casos se ha usado como sinónimo de biodiversidad. Para muchos biólogos, el conteo de especies se ha convertido en una especie de obsesión por cuantificar de alguna manera un fenómeno tan complejo como lo es la diversidad biológica. Un libro reciente sobre el tema, por ejemplo, define a la biodiversidad como “una biología de números y diferencias”.
El Principito no se preocuparía en lo más mínimo si un exobiólogo exageradamente cuantitativo menospreciara su pequeño asteroide debido a su pobreza de especies. Seguramente tal biólogo le haría recordar al hombre de negocios que vivía en otro asteroide y que había logrado contar quinientos millones seiscientos veintidós mil setecientas treinta y una estrellas y que se pensaba dueño de todas ellas por el simple hecho de haberlas catalogado. Podemos imaginar a un exobiólogo visitando el pequeño planetoide del Principito, observando con desdén a su alrededor y comentando en forma despectiva que en la Tierra existe un número aún no determinado de especies de plantas y de animales, pero que la cifra seguramente rebasa las treinta millones de especies. El Principito, sin inmutarse demasiado, se encogería de hombros y preguntaría: “¿Y qué hacen con todas esas especies?”, para rematar con una de sus frases favoritas: “Los adultos son definitivamente extraordinarios”.
Sin duda, la catalogación de las especies de plantas y animales de cada país es un paso importante para el conocimiento y el adecuado uso de la riqueza biológica. Sin embargo, esa labor debe ir aparejada por una comprensión más profunda de lo que implica el término biodiversidad. Hay que recordar que lo que ahora observamos como diversidad biológica es el resultado de millones de años de evolución orgánica y que las diferentes especies que observamos hoy día no son sino un pálido reflejo de la increíble variedad y variabilidad dentro y entre los organismos vivos. Cierto, el conteo de especies nos sirve de guía para estimar la diversidad de un sitio, pero poco nos dice sobre la variación que existe entre los individuos de cada una de las especies, además no toma en cuenta la diversidad genética contenida en esos organismos, ignora la estructura de las comunidades de plantas y animales y olvida la complejidad de las relaciones funcionales dentro de los ecosistemas.
El caso de México es muy ilustrativo. Sin duda es interesante e importante saber que en nuestro país existen alrededor de veinteséis mil especies de plantas vasculares, cerca de doscientos ochenta de anfibios, más de setecientas de reptiles y aproximadamente cuatrocientos cincuenta de mamíferos terrestres. Tal vez también es relevante conocer que nuestro país cuenta con treinta y cinco especies de esquizómidos, diez ricinuleidos, trecerafidiópteros y nueve mecópteros, pero si nos quedamos únicamente con los números fríos y no vemos más allá del simple conteo, caeríamos en el mismo absurdo que el hombre de negocios que se creía dueño de millones de estrellas y lo único que poseía era un papel en el que anotaba sus compilaciones estelares.
Dentro de cada especie, cada individuo es un reservorio único de información genética que se manifiesta en la variabilidad que presentan todas las especies de plantas y animales. Esta variabilidad se hace evidente a nivel local en las diferencias que existen entre los individuos de una población, y a nivel del país en la variación geográfica de las especies. Por ejemplo, todos los venados de cola blanca del país pertenecen a la misma especie (Odocoileus virginianus), pero no se necesita ser especialista en biodiversidad para percibir diferencias entre los venados de un sitio en particular: habrá algunos que tengan las astas más grandes, otros que sean capaces de saltar más alto, o tal vez una de las hembras que tenga un color más pardo que las otras. A nivel geográfico, las diferencias son aún más marcadas: los venados del norte del país son notablemente más grandes que su contraparte en el sureste del país, a pesar de pertenecer a la misma especie. Estas diferencias entre individuos son un componente importante de la biodiversidad que pasamos por alto al simplemente contar especies para cuantificar la riqueza biológica.
En otros niveles de organización existen otras manifestaciones de la biodiversidad. A nivel de las comunidades ecológicas, la composición de especies y las interacciones entre ellas varían considerablemente de un sitio a otro. Para cualquier persona resulta obvia la diferencia entre un desierto y una selva tropical en cuanto al tipo y número de especies de plantas y animales que se encuentran en estos biomas. Sin embargo, la estructura de esos conjuntos de organismos, por ejemplo en cuanto al número de individuos de cada especie y sus caractérísticas morfológicas, resulta menos evidente. Asimismo, la compleja red de interacciones entre los organismos es aún menos conspicua y sin embargo es una parte importante del fenómeno que llamamos biodiversidad.
El asteroide B 612 podrá ser muy pobre en especies, pero su único habitante puede jactarse de conocer y comprender en forma más profunda la biodiversidad de su hogar que cualquier ser humano del planeta Tierra. A través de sus actividades cotidianas, el Principito efectúa un programa eficiente de conservación y manejo de sus recursos. En efecto, al controlar el crecimiento de las plántulas de baobab, regar su preciada flor y deshollinar los tres volcanes del planetoide para evitar erupciones violentas, el habitante de B 612 usa, pero al mismo tiempo protege, la diversidad de este asteroide. Es más, al hacerlo, el pequeño y misterioso ser ama con pasión y se integra en cuerpo y alma a la riqueza biológica de su planeta, por magra que ésta pueda ser.
Desesperado por la codicia del hombre de negocios al contar estrellas, el Principito le espetó: “Es útil para mis volcanes y es útil para mi flor que yo los posea. Pero tú no eres útil a las estrellas”. Este concepto de posesión de la riqueza biológica a través del servicio mutuo es algo que debería hacernos pensar a la hora de diseñar proyectos de manejo de los recursos naturales. Asimismo, deberíamos ir más allá de los simples números, comprender más profundamente la relevancia y las implicaciones de algo tan complicado como lo es la biodiversidad y tratar de ver las miles de sutiles facetas que oculta el fenómeno de la diversidad de los organismos sobre nuestro planeta. Parafraseando al zorro filósofo que se encontró el Principito en la Tierra: la biodiversidad sólo se ve bien con el corazón. Su parte esencial es invisible para los ojos.
|
||||
|
Lecturas adicionales
Saint-Exupéry, A. 1943. Le Petit Prince. Existen muchas traducciones al español de este clásico que nos puede enseñar mucho sobre la biodiversidad y sobre la vida.
Los datos sobre el número de especies en México fueron obtenidos de: conabio. 1998. La diversidad biológica de México: estudio del país. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
|
||||
|
Héctor T. Arita
Instituto de Ecología,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Arita, Héctor T. (1999). La biodiversidad en el Asteroide B 612. Ciencias 53, enero-marzo, 40-42. [En línea] |
||||
 |
 |
||
|
|
|||
|
La creación de la Facultad de Ciencias
|
|||
|
Raúl Domínguez Martínez
conoce más del autor
|
|||
|
|
← Regresar al índice ⁄ artículo siguiente → | ||
|
Es una idea casi unánime que la presencia de México en la vertiente principal de la producción científica y tecnológica a nivel mundial es reconocida como de bajo relieve. De hecho, constituye un lugar bastante común referirse a la inexistencia de una ciencia mexicana (o, mejor dicho, a una ciencia hecha en México, para no ofender a los partidarios de la Ciencia como absoluto), así como a la de una tecnología propia. Desde luego, como todo lugar común, esta afirmación es inexacta y deja de lado una actividad que se ha mantenido vigente aun en contra de obstáculos diversos y de estímulos exiguos y a veces nulos. Trabajos recientes en el campo de la historia de la ciencia están participando en la recuperación de esta faceta del devenir social, que había traspasado los umbrales del olvido al verse cotejada con los momentos estelares de la historia de la ciencia en la civilización occidental, en una comparación de suyo desmesurada.
Los ejemplos abundan respecto de los logros científicos y tecnológicos alcanzados en el país a través del tiempo, aun si nos atenemos a los más conocidos. Ahí está la asombrosa obra de botánica y medicina conocida como Códice Cruz–Badiano, que fue preparada por dos eruditos indígenas en el Colegio de Tlaltelolco para demostrar a Carlos V la capacidad y el nivel de los conocimientos autóctonos en la materia, en muchos sentidos superiores a los europeos de su tiempo; como sabemos, el emperador desestimó esta obra, quedando desde entonces bajo la custodia de la Iglesia en calidad de cosa curiosa. El enorme impulso que recibió la minería a mediados del siglo XVI, se debió al método de amalgamación de la plata con azogue, conocido como “beneficio de patio”, desarrollado en esta tierra por Bartolomé de Medina. La modernización de la astronomía y de la matemática, lograda en el pleno oscuransotismo de raigambre religiosa que prevalecía en la Real y Pontificia Universidad de México durante el siglo XVII, fue merced a las enseñanzas y escritos de un oriundo de nuestros valles centrales llamado fray Diego Rodríguez. La vocación enciclopédica de Alzate y su Diario literario de México así como el caso de otras publicaciones de carácter científico, nos demuestran claramente que la ciencia nunca —menos aún en las sociedades prehispánicas— ha sido algo ajeno a las preocupaciones y prácticas de los mexicanos. Sin embargo, por más minucioso que se haga este recuento de figuras excepcionales, de contribuciones, y aun de periodos en los que se ha configurado una cierta tradición científica, no lograremos encontrar nombres que se pudiesen poner al lado de los grandes artífices de la ciencia occidental. Más aún, no obstante de que contásemos con algún personaje de la talla de Copérnico, Kepler, Curie o Fermi, el problema en realidad se refiere a las condiciones concretas y particulares que han determinado un “subdesarrollo” en materia de ciencia y tecnología, con todas las implicaciones que este término —importado del léxico de la economía política— supone, particularmente, la de dependencia. Es preciso tener en cuenta que el binomio ciencia–tecnología, incluyendo las diferencias históricas y operativas que se puedan reconocer entre estos dos términos, constituye hoy día un factor determinante para definir los niveles reales de desarrollo y de independencia efectiva de una nación. Es decir, una sociedad que en la actualidad no cuente con un aparato sólido y eficaz de producción de ciencia y de tecnología, se encuentra amenazado de una vulnerabilidad extrema que tiende a permear todos los órdenes de la vida colectiva, empezando por el económico. Planteado desde esta perspectiva, el problema adquiere una dimensión distinta y más amplia. La pregunta sería entonces ¿por qué México se ha mantenido a la zaga (lo contrario de la vanguardia) en estos terrenos? Naturalmente, la respuesta es demasiado compleja y me parece que todavía no es posible acceder a ella de manera cabal. Por supuesto, se debe descartar de entrada y en definitiva cualquier recurso explicativo en el que la incapacidad subjetiva juegue algún papel. Semejante precariedad explicativa nos remitiría a la polémica lamentable que sostuvieron algunos destacados pensadores españoles a raíz del descubrimiento de América, como fray Domingo de Betanzos o Pedro Mártir de Anglería, empeñados en dilucidar si los oriundos de estas tierras eran seres humanos o no. La ciencia, así como la tecnología, son efectos de condiciones sociales específicas, en mucho mayor medida que resultado de “genialidades”. El escenario original Al despuntar el presente siglo, la ciencia y la tecnología en México presentaban una panorámica poco alentadora. Los parcos avances que se habían logrado en estos campos, merced a los empeños de los ideólogos porfiristas comprometidos con el prurito cientificista del positivismo, resintieron los estragos de la lucha armada, reduciéndose a una expresión ínfima. La presencia de la tecnología de punta en el país en el contexto del porfiriato no debe incluirse en estas consideraciones, pues hasta los operarios de los equipos eran de nacionalidad extranjera. De hecho, podemos decir que una vez que la Revolución estalló, el de por sí débil aparato de investigación y desarrollo de la ciencia fue desarticulado. Curiosamente, para los efectos del tema que nos ocupa, la única institución porfiriana que habría de sobrevivir a la revuelta sería la Universidad. Como sabemos, esta entidad abrió sus puertas apenas dos meses antes de que, con asombro de puntualidad, la Revolución diese inicio justo en el momento previsto por Madero. Podríamos mencionar, para establecer un contraste paradójico, que el Instituto Tecnológico de Massachusetts (MIT) hizo lo propio el mismo año que dio término la Guerra de secesión en Estados Unidos. Daría la impresión de que la inoportunidad histórica ha marcado nuestros procesos como un sino. El caso es que mientas el vértigo de la violencia iba dando cuenta de lo poco —o mucho, según el parámetro que se quiera emplear— que se había logrado estructurar para fomento de la ciencia, la flamante y amenazada Universidad Nacional se aferraba con hálito vital a la existencia que, gracias a la convicción y a los desempeños de hombres insignes, ha conservado hasta la fecha. Ahí se sostuvo el impulso en favor de la ciencia, pero no logró salir inerme; ya que la Escuela de Altos Estudios, vanguardia indiscutida del desarrollo científico del país y parte constitutiva de la Universidad Nacional por decisión ejecutiva, pasó de ser una institución predominantemente científica a humanística, llegando, en el transcurso de 1912 y a consecuencia de los drásticos recortes presupuestales ¡ojo!, al extremo de verse incapacitada incluso para la contratación de profesores mexicanos regulares, siendo que hasta entonces contaba dentro de su planta con prestigiados académicos extranjeros. Es de suponerse que el costo menor de las indagaciones humanísticas fue lo que marcó la pauta para que lo científico cediera terreno. La nueva orientación de Altos Estudios se perfiló, en lo que a ciencia se refiere, hacia la creación de cursos libres, llamados así porque no dependían ni de financiamiento, ni de matrícula, ni de sistematización formal, ni de nada, como no fuera la vocación y el ánimo inquebrantable de unos cuantos eruditos empeñados en que la sociedad contara con espacios para la actividad científica. Adolfo Castañares, en Química, y Sotero Prieto, en Matemáticas, son dos de los conspicuos personajes que mantuvieron vivo el interés por la ciencia en circunstancias muy poco propicias. Sin embargo, para darnos una idea de lo que esto representaba a nivel nacional, diremos que en esas fechas, Sotero Prieto impartía sus cursos a un total aproximado de veinticinco estudiantes. ¿Qué sentido guardaban estas proporciones en una sociedad que se abría paso en pleno siglo XX? Muy pobre, sin duda, pero superior a nada. Tengamos en cuenta que antes de que se iniciara la Revolución en México ya se había logrado la comunicación telegráfica sin hilos a través del Atlántico; Orville Wright había conseguido el vuelo de un artefacto propulsado, más pesado que el aire y ya habían aparecido los trabajos de Lorentz, Einstein y Minkowski sobre la relatividad restringida. La ciencia y la tecnología se encontraban en auge y su papel en el desarrollo de los pueblos adquiría un relieve cada vez más acusado; lejos quedaban los tiempos en los que el conocimiento científico había suministrado la máquina de vapor a una industria basada casi por entero en las técnicas tradicionales y que debía mucho más al talento de los artesanos que a la ciencia. Ahora, la ciencia asumía un papel protagónico y estrechaba nexos con la tecnología. Si se me permite la expresión, diría que el siglo XX comenzaba para Occidente bajo el signo de tecnologías con una cuota creciente de sofisticación científica incorporada. Era —y sigue siendo— la pauta del progreso. Y la nación mexicana entró al siglo soslayando esta promisoria vertiente. En 1917 se reinició la vida constitucional en México. La turbulencia bélica quedaba atrás y se daba comienzo al proyecto nacional, que, según constatamos a la distancia, no fue tan distinto del porfiriano. La educación fue enarbolada como parte fundamental del proyecto, con un marcado sesgo populista que desde luego hizo de lado la atención a la educación superior. La ciencia no formó —como no forma hasta ahora— parte del proyecto nacional. El aparato educativo fue descentralizado y se transformó en administración municipal, desapareciendo la Secretaría de Instrucción Pública. Se creó, en cambio, el Departamento Universitario y de Bellas Artes con el presupuesto más bajo de todas las dependencias gubernamentales: ese año de 1917, se le asignaron cuatro millones de pesos, frente a los ciento veinte millones que recibió la Secretaría de Guerra. El impacto se resintió aun después de concluida la etapa militar: en 1910, el presupuesto ordinario destinado a educación, todavía bajo la administración de Díaz, representaba 7.0% del total; en 1917 esta relación fue de 1.2% y de 0.8% al año siguiente, debiendo esperar hasta 1922 para que el gasto educativo mejorase su participación relativa, con un 12.9% sobre el total. Lo anterior obedecía claramente a una realidad; de acuerdo con los datos del III Censo General de Población aplicado en 1910, en México había 15 160 000 habitantes, de los cuales poco menos de medio millón se ubicaba en la ciudad capital. La población rural, en comunidades menores de diez mil habitantes, representaba 86.5% del total, y la población analfabeta, con diez años o más, ascendía a 72.3%. Es evidente que el objetivo prioritario, y urgente, era el de dar atención a segmentos muy amplios de la población que carecían de enseñanza básica, subordinando y postergando el fomento a la educación superior y, con ella, al desarrollo científico. De esta manera, México dio comienzo a la fase contemporánea de su historia, con la virtual inexistencia de un aparato científico y tecnológico y con la disponibilidad de fomentar estos campos, reducida a una sola instancia: la Universidad Nacional. El papel estratégico de la universidad Las naciones que han marcado la pauta del progreso científico y tecnológico en el mundo, y de manera particular durante el presente siglo, han contado —piénsese en cualquiera de las que integran el con-junto— con un aparato bien estructurado que se encarga de gestar el desarrollo en tales campos y que, además de ser funcional, se encuentra articulado con otros componentes de la realidad social. Los dispositivos que conforman este aparato son de índole diversa e incluyen universidades, tecnológicos, academias de ciencias, laboratorios, centros de investigación privados y públicos, y algunos otros más específicos. Es evidente que en todos los casos se trata de una responsabilidad compartida y de funciones diferenciadas. En el caso de México, esto no ha sido así. De hecho, la gran mayoría de estas actividades han sido confiadas a una única institución: la Universidad Nacional. Sólo hasta fechas más o menos recientes esta ingente responsabilidad se ha ido descargando y compartiendo con otras instituciones. Podemos decir, sin exageración, que los dos primeros tercios del presente siglo, la así llamada Máxima Casa de Estudios del país, con la honrosa y excepcional colaboración de otras instituciones de mucho menor cobertura como el IPN, ha encarado por su cuenta estos amplísimos cometidos, supliendo el lugar y las funciones de entidades que, por inexistencia (las universidades y tecnológicos que el desarrollo de la sociedad ha reclamado), por ineficiencia (todos los organis mos antecesores del CONACYT) o por insuficiencia, han generado un vacío. Sus aportaciones en el campo de la investigación han sido —sencillamente— imprescindibles. No existe duda alguna de que la mayor parte de lo que en México se ha logrado en materia de ciencia, en un doble sentido cualitativo y cuantitativo, ha tenido su origen en la Universidad Nacional. Las anteriores consideraciones, aparte de ser un justo reconocimiento, sirven para abordar de manera analítica la función de la Universidad en el contexto de la República. Se trata de un papel estratégico que ha rebasado de forma franca los linderos de una institución particular, para ubicarse en una perspectiva mucho más amplia. Los mexicanos estamos de cierto familiarizados con formas cuyos contenidos efectivos no se corresponden con las definiciones nominales. ¿Cómo explicar, por ejemplo, la existencia de una República democrática y federal en donde un sólo partido ha monopolizado el poder por setenta años y todas las decisiones se toman desde el centro? El asunto se refiere en realidad al funcionamiento de determinados mecanismos o dispositivos que operan a manera de subterfugio, con los cuales se ejerce una práctica que es distinta a la que se designa en la definición formal correspondiente. En el caso que nos ocupa, el de la Universidad, este subterfugio ha tenido como eje dos elementos operativos: el concepto de autonomía y la categoría de nacional. De estos dos elementos se ha valido el poder público para realizar una transferencia de responsabilidades, sin la correspondiente transferencia de recursos efectivos y suficientes, lo que le ha permitido desentenderse directamente de tales responsabilidades, a la vez que mantener su imagen como patrocinador. Es decir, hacia ella se fueron canalizado obligaciones que no han sido acompañadas de dotaciones pecuniarias adecuadas, dejando la respuesta a las limitadas posibilidades de la institución y desbordando el ámbito propio de su competencia. En este horizonte, la Universidad se ha visto compelida a la formación de profesionales en ramas descuidadas por otras instituciones en cuanto a la calidad y también a la cantidad; a la creación de planteles de enseñanza y de investigación que el desarrollo o las previsiones de desarrollo reclaman, y que no son atendidas por alguna otra instancia; a la realización de investigaciones “sobre los problemas nacionales,” financiando, formando investigadores, procurando equipos y laboratorios, etcétera, abarcando un espectro en el que potencialmente todo queda incluido; a la formación de la planta académica de otras instituciones; a la colaboración en el trabajo de elaborar planes de estudio en dependencias ajenas; a la impartición de cursos especiales; a la creación de especialidades y posgrados; al otorgamiento de becas; a la difusión del conocimiento; etcétera, etcétera. Así, podemos afirmar que gran parte de lo que en materia de ciencia y de tecnología se ha realizado en el país se le debe a la Universidad Nacional y que ésta ha actuado de conformidad con su propia vocación, su propia inercia y con recursos escatimados que nunca resultaron suficientes y que ha tenido que administrar con malabarismos que muchas veces rayan en lo milagroso. Más adelante veremos cómo en el caso particular de la Facultad de Ciencias, así como en el de algunos institutos como el de Matemáticas o el de Física, su existencia fue resultado de iniciativas que la Universidad Nacional emprendió por cuenta propia y de las cuales no existían —y no lo hubo por bastante tiempo— equivalentes en todo lo largo y ancho del territorio nacional. Antes de cerrar este apartado, sería conveniente añadir una precisión de gran importancia para ubicar correctamente lo que se ha expuesto. Aparte de la enorme negligencia con la que el Estado mexicano actúa cuando se trata de asuntos que no le son rentables políticamente en un plazo inmediato, la cuestión del escaso desarrollo de las ciencias en nuestro país, problema de características crónicas, se relaciona más a profundidad con el desarrollo de las fuerzas productivas. La producción de ciencia y de tecnología propias no se ha presentado históricamente en nuestro país como una necesidad generada por la sociedad, la cual ha satisfecho esos requerimientos por la vía de la importación. Una sociedad atrasada, marcada por la desigualdad, y una economía desequilibrada, marcada por la dependencia, constituyen condiciones de fondo que se reproducen a sí mismas, determinando un rango de factibilidad sumamente restringido para la ciencia generada en el país. La ciencia y, desde luego, el desarrollo tecnológico resultan inversiones rentables a mediano plazo; el problema es el del financiamiento a corto plazo. Tengamos en cuenta que la burguesía doméstica no está “acostumbrada” a invertir en negocios que no le reditúen ganancias inmediatas y seguras. Se trata de una burguesía local marcada asimismo por el atraso, con una jerarquía de intereses en donde los nacionales se subordinan. No ha resultado, pues, un agente viable para el fomento a la ciencia y la tecnología, como se ha constatado en los hechos; la alternativa, en semejantes condiciones, quedó a cargo del Estado, el que, a su vez, se sirvió delegar esa tarea a un reducidícimo grupo de instituciones encabezadas por la UNAM. La autonomía y el aislamiento A pesar de la acusada desconfianza que los gobiernos emanados de la Revolución le profesaban a la Universidad Nacional, ésta logró sobrevivir y aun incrementar su presencia en el marco de la sociedad. Fueron conflictos ya ajenos a su raigambre porfiriana los que produjeron nuevos focos de conflicto entre la Universidad y el poder público. En efecto, después de la Revolución se pueden identificar tres etapas distintas (hoy día parece configurarse la cuarta) en las cuales la Universidad Nacional se ha redefinido en función de la posición que guarda en el esquema de la administración pública: en 1929 se le concedió una autonomía restringida, con la correspondiente promulgación de una Ley Orgánica; en 1933 se le concedió una autonomía total y se suprimió su carácter de nacional, quedando a expensas de un subsidio aleatorio; en 1945, por convenir así a intereses emergentes, el poder público retomó la tutela de la institución, promulgando la Ley Orgánica vigente. Hay que mencionar que no obstante estos drásticos cambios, la Universidad prosiguió atendiendo sus compromisos de manera ininterrumpida. En 1933, la Universidad tenía matriculados a poco más de nueve mil alum-nos en todos sus planteles, incluida la Escuela Nacional Preparatoria, con un crecimiento relativo de su población del orden de 13% respecto de la inscripción de 1929. En el mismo año, se estima que la población total de la República Mexicana era ligeramente superior a los diecisiete millones, lo que significa que la ya desde entonces Máxima Casa de Estudios del país atendía apenas a 0.05 % del total, cifra insignificante en términos absolutos, máxime si se tiene en cuenta que de tal cantidad sólo una porción reducida alcanzaba la titulación. Por otro lado, estos totales acusaban un fuerte desequilibrio en favor de las “carreras liberales”: las escuelas de Jurisprudencia y de Medicina absorbían, ellas solas, a 38.7 % de la matrícula completa de nivel superior. Como se puede inferir con éstos y otros indicadores, el aporte de la Universidad al desarrollo del país en ese tiempo era, por decirlo de alguna manera, irrelevante. Si ésta era la situación con las carreras más populares, se puede uno imaginar lo que ocurría con las ciencias; baste por ahora con decir que las carreras de Biología, Matemáticas o Física se impartían en la Facultad de Filosofía y Letras, y que tenían un perfil por completo diferente al que ahora tienen. Las de Biología, Matemáticas y Física eran carreras que se impartían a la manera de una Escuela Normal Superior, es decir, para preparar profesores en tales disciplinas. A lo anterior habría que añadir que para la nación —o, mejor dicho, para el programa enarbolado por el gobierno federal, que en el caso nuestro sabemos que no es lo mismo— la presencia de una Universidad insumisa venía siendo, además de poco menos que suntuaria, incómoda. Ello quedó demostrado de manera fehaciente con la promulgación de la Ley Orgánica de 1933, cuando le fue concedida la autonomía total, desentendiéndose el poder público de todo tipo de apoyo, incluido el pecuniario; así quedó asentado en el inciso II, apartado b) del artículo 9 de la Ley aprobada por el Congreso de la Unión en el mes de octubre: “Cubiertos los diez millones de pesos (que el gobierno federal entregará a la Institución), la Universidad no recibirá más ayuda económica”. Las palabras del licenciado Narciso Bassols, entonces secretario de Educación Pública, al ser discutida la iniciativa de ley a la que me refiero, corroboran lo dicho: “no se puede decir que la Universidad haya realizado con provecho sus destinos; no se puede decir que la acción educativa haya progresado. La Universidad tiene una enorme, una grave responsabilidad ante la República, y sólo porque la masa de habitantes del país, situada más allá de la ciudad, no puede apreciar y sentir de un modo palpitante e inmediato, no puede conocer las intimidades de las deficiencias universitarias, no se ha producido una poderosa y tremenda reacción de protesta nacional”. La ruptura con el poder público habría de reportarle a la Universidad una situación de fuerte insolvencia. Pero más allá de esta clase de diferencias entre ambos, lo que realmente sitúa en perspectiva correcta el desarrollo de la institución y su papel en el México de la época, es lo que se refiere a la necesidad de la sociedad en general, y de la planta productiva en particular, de los servicios que ésta ofrecía o podía ofrecer. Es preciso tener en cuenta en este aspecto que el México de los años treintas era un país fundamentalmente agrario. En 1930, 80.7% de los mexicanos vivía disperso en localidades menores de diez mil habitantes, es decir, rurales; el sector primario absorbía a 3 626 trabajadores (PEA) sobre un total de 5 151, es decir, a 70.3% . Y si bien la porción mayoritaria del Producto Interno Bruto no tenía su origen en este sector, debido a su baja productividad (cinco veces menor a la productividad del sector industrial, por ejemplo), la producción agropecuaria y minera constituía la base de la actividad exportadora, determinando de tal suerte que el país estuviese convertido en un importador neto de bienes manufacturados. La tasa de analfabetismo rebasaba el 60% de la población con diez años de edad o más, en el seno de una sociedad polarizada en donde los estratos medios eran minoría. Parece claro que la institución universitaria no era, llanamente, una prioridad nacional. Su relevancia a nivel nacional era ínfima, tanto en lo cuantitativo como en lo cualitativo. Por ello, lo que en realidad significó para el poder público la concesión de la autonomía total, fue la posibilidad de desembarazarse de un problema y de desentenderse de un cometido que, en la forma, se debería cumplir. La línea política que adoptó el general Cárdenas al asumir el gobierno de la República en noviembre de 1934, vino a reforzar más esta situación. En el texto de una carta fechada en septiembre de 1935 para dar respuesta a una solicitud del rector procurando el apoyo económico del gobierno, el presidente de la República acusa a la Universidad de haberse colocado “por su propia voluntad, en un plano de indiferencia con respecto al Programa Social de la Revolución”. Poco tiempo más tarde, en 1937, abrió sus puertas el Instituto Politécnico Nacional, concebido y planeado como una alternativa al liberalismo universitario, dotado de mecanismos tales que el Estado aseguraba un amplio radio de control sobre sus procesos. Frente a semejantes condiciones, los desempeños de la institución educativa se vieron amenazados con severidad. Hacia fines de noviembre de 1937, el rector Chico Goerne preparó una detallada exposición de la situación por la que ésta atravesaba, con objeto de procurar fondos federales. En ella señalaba, por ejemplo, que los fondos solicitados para evitar un desgaste más acentuado no representaban ni 1% de los egresos del erario; que los honorarios del profesorado se encontraban a niveles inferiores a los devengados diez años antes, ya de suyos exiguos, encontrándose entre los más bajos del mundo, y que los profesores debían atender normalmente a grupos no menores de cien alumnos y, en muchas ocasiones, superiores a trescientos. En ese tiempo, las reducciones voluntarias de salario se volvieron más o menos frecuentes, las donaciones de equipo constituyeron una alternativa ante la incapacidad de la institución de allegárselo por sí misma, llegando al grado de que en 1940 el Banco de México hubo de donar mil sillas para poder sentar a otros tantos alumnos a recibir sus clases. Las obras de infraestructura quedaron congeladas e, incluso, diversos bienes inmuebles empezaron a evidenciar un fuerte deterioro por falta de mantenimiento. En fin, se trataba de una Universidad con un muy marcado déficit económico, objeto de una cierta animadversión por parte del Gobierno Federal y con una presencia sumamente débil en la sociedad, convulsionada por desordenes internos recurrentes y sujeta a una estructura de gobierno que, sobre la base de la paridad, no facilitaba hacer prevalecer los criterios académicos en la toma de decisiones. El estado y la evolución de la ciencia La Escuela de Altos Estudios fue disuelta en 1925 para dar paso a la creación de la Facultad de Filosofía y Letras. Hasta ese momento, los cursos y las investigaciones relativas a las ciencias habían tendido a la desaparición de una filiación institucional, convertidos en acciones de personas singulares que no estaban —no podían estarlo— sujetas a una normativa, ni a una sistematización, ni a la supervisión colegiada, ni mucho menos a una regulación por concepto de emolumentos, tal y como ocurre ahora o como ocurría ya entonces en universidades de mayor desarrollo fuera del país. Si bien es cierto que por la cuota de entusiasmo que ponían los responsables de estas actividades académicas se logró inculcar interés por la ciencia en algunos jóvenes estudiantes que más tarde se convirtieron en científicos de gran prestigio, incluso internacional —ahí está el caso de Sandoval Vallarta, por ejemplo—, la perspectiva y el nivel en el que se desenvolvían eran considerablemente más bajos. En realidad, se trataba casi en exclusiva de actividades de cultura general y, en el mejor de los casos, de formación de profesores. Es evidente que en esa época no existía en todo el país disponibilidad de empleo para que egresados de matemáticas o de física se desempeñaran como tales. La herencia con la que la institución se acercaba a su vida autónoma era ciertamente pobre. En matemáticas, la Escuela de Altos Estudios había podido ofrecer tan sólo cuatro cursos libres, sin ningún plan de estudios que les diera unidad, a cargo de Sotero Prieto, Juan Mancilla y Río, Luis Espino y Daniel Castañeda. En Física se impartieron únicamente dos cursos entre 1912 y 1914, a cargo de Valentín Gama y Joaquín Gallo. La rama de Química fue más favorecida y en 1916 se fundó la Escuela de Ciencias Químicas, a cargo del ingeniero Salvador Agraz. En Biología se llegó incluso a la impartición de dos carreras: la de profesor en Botánica y la de profesor en Zoología; éstas se fundieron en una sola a partir de 1922. Sin embargo, al ocurrir en 1925 la conversión en la Facultad de Filosofía y Letras no se registró ninguna repercusión en el campo de las ciencias, las cuales prosiguieron con su ritmo habitual. El paso siguiente ocurrió hasta 1928, cuando se reformaron los planes de estudios de las diversas carreras, procurando elevar los objetivos y el nivel académico. La marcha continuaba con paso en exceso lento. No obstante, entonces se gestaba un elemento importante que pronto rendiría frutos positivos, relativo a que ya se había iniciado el envío de estudiantes mexicanos al exterior. En el informe del rector ante el Consejo Universitario, presentado por Ignacio García Téllez, se encuentra una parte en donde dice: “[La Universidad] llama a su seno a los mexicanos que estudian en el extranjero, expuestos a perderse como factores de integración patria, y entre tanto regresan, procura estar en contacto con ellos, enterándolos de nuestra vida y recordándoles su país”. Desde luego, hay que tener en cuenta que estos estudiantes en el extranjero, en particular los de ciencias, tenían ante sí una disyuntiva complicada por obvia: o permanecían en el exterior, o renunciaban a su formación específica, involucrándose en la-bores un tanto ajenas a ella, o se incorporaban a la Universidad como personal académico, ya que ésta era la única instancia en el país con características capaces de asimilarlos en un plano profesional. Ellos habrían de conferirle un sesgo diferente y cualitativamente superior a las ciencias enseñadas e investigadas en la Universidad, la que, por su parte, no cejó jamás de escudriñar y de aprovechar toda circunstancia que le redituara una mejoría.
En 1931, la Facultad de Filosofía y Letras se encontraba dividida en cuatro secciones: Filosofía, Letras, Ciencias Históricas y Ciencias. Los grados que en esta última sección se concedían eran los de maestro y doctor en Ciencias Exactas; maestro en Ciencias Físicas, y maestro y doctor en Ciencias Biológicas. Dicha Facultad estaba ubicada en la calle San Ildefonso número 33, en el centro de la ciudad. Entonces, la población escolar de la Universidad ascendía a casi diez mil alumnos, de los cuales únicamente trescientos sesenta y nueve estaban inscritos en todas las carreras que esta Facultad impartía. Una reorganización general emprendida en la Universidad en 1935, dio lugar a la creación de una Escuela de Ciencias Físicas y Matemáticas. Con el principio de agrupar áreas académicas en torno de unidades docentes que fungirían a manera de ejes, esta Escuela emergió como parte integrante de la Facultad de Ingeniería y su misión principal, al menos de acuerdo con la visión de su principal promotor, el ingeniero Monges López, sería la de preparar futuros investigadores. La sede que le dio albergue fue el Palacio de Minería. Este mismo año ocurrió el trágico deceso de Sotero Prieto, pilar fundamental del desarrollo de las matemáticas y aun de la física en nuestro país. En diciembre de 1938 se promulgó un nuevo Estatuto General para la Universidad, con el cual comenzó de manera formal la existencia institucional de la Facultad de Ciencias. Al año siguiente, en el que propiamente iniciaron sus funciones, y de acuerdo con datos de la Secretaría General, la nueva Facultad contó con una matrícula de ciento catorce alumnos, de ellos cuarenta y seis eran hombres y sesenta y ocho, mujeres. Hasta entonces se habían expedido exclusivamente títulos de maestría en ciencias biológicas y uno sólo de profesor en matemáticas, concedido en 1938. La inmensa mayoría de los que ejercían como titulares ostentaban en realidad el título de ingeniero, pero a partir de ese momento se iniciaba una etapa completamente distinta en beneficio de las ciencias en la Universidad y, por ende, en México. El momento de la creación En octubre de 1938 el Consejo Universitario recibió un proyecto relativo a la creación de la Facultad de Ciencias; se trataba de un documento de primera importancia, no sólo porque constituía el origen de dicho plantel, sino porque en él se exponían los motivos por los que se tomó tal decisión. Dicho proyecto lo suscribieron el doctor Antonio Caso, director de la Facultad de Filosofía y Estudios Superiores; el doctor Isaac Ochoterena, director del Instituto de Biología; el ingeniero Ricardo Monges López, responsable de la Escuela Nacional de Matemáticas y Ciencias Físicas, y el doctor Alfredo Baños, director del Instituto de Ciencias Físico-Matemáticas. Los argumentos daban inicio con esta afirmación: “En todas las principales universidades del mundo, aun en las de segundo orden, existe una Escuela o Facultad dedicada al estudio superior de las ciencias”. Acto seguido, se deslindaba la enseñanza y el estudio de las ciencias en la universidad como elementos de cultura general, de la enseñanza y el estudio especializados, para después pasar a una crítica de la situación particular: “En nuestro medio universitario se ha tenido especial cuidado de formar buenas escuelas profesionales, dotándolas, hasta donde se ha podido, de todo lo que necesitan para preparar a sus alumnos, pero a los profesores universitarios y a los investigadores de la ciencia no se les ha prestado ayuda alguna, se han formado por su propia cuenta”. De conformidad con los propios autores, tal situación “no ha sido por falta de esfuerzos encaminados a ese fin, sino porque en los últimos años hemos vivido largos periodos de inquietud y hemos sufrido una continua falta de recursos que ha obligado a nuestras autoridades universitarias a posponer la resolución de los problemas culturales para atender a la imperativa necesidad de subsistir”. La propuesta consideraba “cuando menos” siete departamentos: Matemáticas, Física, Química, Biología, Geología, Geografía y Astronomía, y la creación consecuente de otros tantos institutos de investigación, razón por la cual “se propone que el Instituto de Ciencias Físico-Matemáticas se divida en dos institutos, uno dedicado a las matemáticas y otro a la física; que se establezca el Instituto de Química, que tanta falta hace, y que el Instituto de Investigaciones Geográficas se incorpore con el nombre de Instituto de Geografía […] de modo tal que cada jefe de instituto será al mismo tiempo, exoficio, jefe del departamento respectivo de la Facultad de Ciencias”. No deja de llamar la atención, por último, el que los términos de la propuesta se hallan limitado, como era lo usual, a reformulaciones legales, sin acompañar la exposición de motivos de una consideración relativa a los recursos humanos y físicos, y a la relación entre lo disponible y lo deseable. La organización, el reglamento y los planes de estudio para la nueva facultad fueron elaborados, en las ramas de física y matemáticas, por el ingeniero Monges López y por el doctor Alfredo Baños, quien recién había regresado al país después de graduarse en la Universidad John Hopkins y de haber recibido la beca Guggenheim, siendo el primer doctor en Física mexicano con residencia aquí (Sandoval Vallarta vivía en Estados Unidos). El documento fue presentado a la consideración del Consejo Universitario en noviembre de 1938. La iniciativa establecía diversos mecanismos de coordinación entre los departamentos y los institutos respectivos, esto con la clara intención de dar un fuerte impulso a la investigación y de integrarla al trabajo docente, razón por la cual el documento incluyó la propuesta de dividir el Instituto de Ciencias Físico-Matemáticas, de reciente creación, en dos institutos especializados, dando lugar así al surgimiento de un Instituto de Física propiamente dicho; el de Matemáticas sería posterior. La nueva Facultad otorgaría los grados de maestro y doctor en Ciencias, con una nomenclatura peculiar compartida con Filosofía y Letras en la que la maestría era equivalente a la licenciatura, y siendo requisito la obtención del primero para poder cursar el doctorado. El 19 de diciembre de 1938 se promulgó el nuevo Estatuto General; de esta manera dio comienzo la vida formal tanto de la Facultad de Ciencias como del Instituto de Física. El artículo 8 establecía que “los institutos tendrán un director, constarán de las secciones con el personal técnico y administrativo que señale el reglamento y dependerán directamente del rector. Cada uno de los institutos tendrá un Consejo consultivo [antecedente del actual Consejo Técnico de la Investigación Científica] que deberá hacer al rector todas las observaciones que estime pertinentes para el mejor desarrollo de los trabajos”. Los directores de institutos podrían durar indefinidamente en el cargo, de acuerdo con el artículo 34, pudiendo ser removidos por el Consejo a solicitud del rector o de un grupo de consejeros que representaran cuando menos un tercio de los votos computables en el órgano legislativo. Las labores efectivas dieron inicio en enero de 1939, año para el cual había sido aprobado el Reglamento de Pagos, estableciendo una cuota de inscripción para los alumnos de la Universidad de diez pesos y una colegiatura anual de cien pesos para los de la Facultad de Ciencias; ese año, después de una devaluación brusca, el dólar se cotizó a $5.19, y el salario mínimo diario general en la zona metropolitana era de $2.50, lo que suponía, para los alumnos de física, un pago anual equivalente a poco menos de veinte dólares y a cuarenta salarios mínimos. Adelantaremos aquí que, para 1940, la matrícula de la nueva Facultad representaba sólo 1.1 % de la población estudiantil universitaria. Para el caso de la física y la matemática, la Facultad de Ciencias continuó ocupando las instalaciones en el Palacio de Minería, y el edificio porfiriano de Ezequiel Montes número 15 albergó el área de la biología. El cambio más importante era, en realidad, el de perspectiva, ya que se había logrado el paso a la creación de una entidad autónoma para el desarrollo de las ciencias. El énfasis inicial fue puesto en el desarrollo de las matemáticas y la física y en la preparación de cuadros en el extranjero. Por su parte, el Instituto de Física comenzó sus trabajos en medio de dificultades principalmente de orden pecuniario. El primer ejercicio presupuestal no pudo hacerse efectivo debido a problemas administrativos; esa primera asignación fue de veinte mil pesos anuales, cifra que se elevó a treinta y ocho mil pesos en 1939. Aquí conviene recordar que en el curso de ese año dio comienzo la conflagración bélica en Europa, en cuyo contexto habría de tener lugar un impresionante desarrollo de la física nuclear. Para entonces, el Instituto ocupó un local dentro del Palacio de Minería, en la calle de Tacuba, en el centro de la ciudad. Aparte del doctor Baños, director del mismo y precursor en México, junto con Sotero Prieto y Blas Cabrera, de los estudios de Física Atómica y de Física Teórica, figuraban como investigadores el ingeniero Manuel González Flores y los profesores (aún no contaban con el título profesional) Héctor Uribe y Manuel Perusquía, así como un ayudante de investigador, el señor Pedro Zuloaga. Radiación Cósmica, Hidrodinámica y Elasticidad, Física Biológica, Física Nuclear y Radioactividad, Espectroscopía y Estructura Atómica, Rayos X y Estructura Molecular, Astrofísica, Geofísica y Laboratorio de Mecánica de Suelos eran las secciones que integraban el núcleo estructural del Instituto, según su plan original. Sin embargo, dadas las limitaciones presupuestales y, en general, las carencias de la Universidad, hubo de conformarse con establecer y hacer funcionar las secciones de Radiación Cósmica y de Mecánica de Suelos, e iniciar los trabajos teóricos de la geofísica. En un artículo preparado por el doctor Baños a finales de 1940 para la Revista de Estudios Universitarios, órgano de difusión de las Facultades de Filosofía y Letras, Ciencias y de la Escuela Nacional Preparatoria, el director del Instituto presentó a la comunidad universitaria las siguientes reflexiones: “A diferencia de otros institutos, tales como el Instituto de Biología, el Instituto de Geología y el Observatorio Astronómico Nacional, que ingresaron a la Universidad Nacional Autónoma de México en 1929, dotados de amplio patrimonio en cuanto a edificios, laboratorios, bibliotecas y personal, el Instituto de Física ha empezado sin patrimonio alguno, con lo cual la labor de creación a la vez que de organización y dirección se torna de lo más difícil, especialmente en vista de las circunstancias precarias por las que ha atravesado la Universidad durante los últimos tres años escasos que lleva de vida el Instituto de Física. Sin embargo, es la esperanza de esta Dirección que las autoridades universitarias, animadas de un alto espíritu de deber universitario y percatadas por entero de la importancia y trascendencia de la misión que el Instituto de Física está llamado a desempeñar dentro de la Universidad como laboratorio central de pruebas y medidas físicas y como centro de investigaciones en las ciencias físico-matemáticas, habrán de prestarnos su más amplia colaboración y franca ayuda para dotar, desde luego al Instituto, aunque sea de forma modesta y sencilla, de un edificio adecuado para sus oficinas, sus laboratorios y su biblioteca”. Estos acontecimientos fueron reconocidos por el rector Gustavo Baz en el informe que presentó en 1940: “De enorme trascendencia para el país fue la creación de la Facultad de Ciencias. La Universidad de México estaba integrada por un conjunto de Escuelas en las que se preparaba a los profesionistas. Pero nos faltaba la pura investigación científica y particularmente la preparación de los investigadores, hombres ajenos al ejercicio práctico de una profesión y que deben tener como fin la pura investigación científica”. Así fue como se inició en nuestro país la promoción institucionalizada de científicos, con presupuestos exiguos y escatimados, en un contexto generalizado de apatía, sin conexiones operativas con la planta productiva, y careciendo de estímulos sociales en favor del incremento de vocaciones en las diversas áreas. |
|||
|
Raúl Domínguez Martínez
Centro de Estudios sobre la Universidad,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Domínguez Martínez, Raúl. (1999). La creación de la Facultad de Ciencias. Ciencias 53, enero-marzo, 4-13. [En línea] |
|||
| ← Regresar al índice ⁄ artículo siguiente → |
| revista de cultura científica de la Facultad de Ciencias de la Universidad Nacional Autónoma de México |
| portada articulo 1 | portada articulo 1 | ||
|
|
|||
|
Genes ¿para qué?
|
|||
|
Helène Gilgenkrantz, Jacques Emmanuel Guidotti
y Axel Kahn.
conoce más del autor
|
|||
|
|
← Regresar al índice ⁄ artículo siguiente → | ||
|
Nuestra percepción actual acerca de la genética está sufriendo cambios profundos. Desde los primeros trabajos de Gregorio Mendel en 1865, hasta el desarrollo de la enzimología y de la bioquímica de las proteínas en las décadas de los cincuentas y los sesentas, los estudios de genética se basaron en el análisis de la transmisión de caracteres fenotípicos de una generación a la otra, es decir, en el estudio de los caracteres observables. A partir de 1973, con el inicio de la ingeniería genética, se empezó a considerar que el procedimiento normal para identificar un gen consistía en caracterizar primero la proteína para la cual codifica. A mediados de los años ochentas, armados con la experiencia de la ingeniería genética y de los estudios de adn, se emprendió un retorno a las raíces: se estableció que el gen responsable de una enfermedad se puede identificar porque cosegrega (se transmite) con un fenotipo patológico, lo que se conoce como clonación posicional. La proteína se deduce a partir de la secuencia del gen identificado, e incluso se puede sintetizar por ingeniería genética. Este procedimiento también permite reconocer genes cuyas mutaciones no son directamente responsables de una enfermedad, pero que sin embargo aumentan la probabilidad de que ésta se manifieste, como en el caso de los genes de susceptibilidad a la arteriosclerosis, hipertensión o Alzheimer.
Recientemente la sed por descifrar nuestro propio genoma nos ha llevado a una nueva era, la de la genómica, cuyo principal objetivo es la secuenciación completa del genoma humano. De hecho, acaba de anunciarse la primera secuencia entera de un cromosoma humano, y podemos asegurar, sin demasiado riesgo, que todos los cromosomas habrán sido secuenciados en un futuro muy próximo.
Sin embargo, aún nos falta un enorme trecho antes de que a partir de la secuencia de los genes podamos inferir los fenómenos fisiológicos y patológicos. Entre los ochenta mil a ciento cuarenta mil genes de los que se compone nuestro alfabeto, sólo algunos están implicados en una enfermedad monogénica específica. Algunos de ellos son tan importantes que la más ligera mutación es letal, pero en otros casos existen candados de seguridad que permiten a ciertos genes paliar la ausencia o la alteración de otros, por medio de una cierta redundancia funcional. Además, el resultado fenotípico de un gen depende, generalmente, de su colaboración con otros genes y del contexto del medio ambiente, de la misma forma que el significado de una palabra depende de su posición dentro de una frase y debe de ser interpretado en función de la trama de la historia.
Ratones para los hombres
Uno de los prerrequisitos indispensables para la realización de ensayos terapéuticos clínicos es contar con modelos animales. A partir del nacimiento de la transgénesis, que permite la introducción de un fragmento de adn en el genoma desde los primeros estados de la embriogénesis, ha sido posible desarrollar diversos modelos de afecciones humanas. Por medio de esta técnica no sólo se pueden modelar enfermedades monogénicas, sino que también se han desarrollado modelos de cáncer en tejidos específicos y es posible estudiar la cooperación entre distintos oncogenes, la cinética de la aparición de un proceso canceroso y, de esta manera, establecer sitios potenciales de intervención terapéutica.
Hacia finales de la década de los ochentas se desarrolló una herramienta de creación de modelos animales muy poderosa; se trata de la posibilidad de reemplazar, durante las primeras etapas del desarrollo embrionario de los ratones, un gen normal por uno mutado, lo que se conoce como recombinación homóloga. Hasta entonces los investigadores habían dependido del carácter aleatorio de la mutagénesis, pero con esta nueva técnica, que se basa en el conocimiento del gen que se quiere interrumpir o mutar, se introduce la secuencia mutada en células embrionarias totipotenciales de ratón y, por medio de un método de selección positiva o negativa, se escogen aquellas células donde la secuencia mutada haya reemplazado a la secuencia normal del ratón. Gracias a esta técnica ha sido posible desarrollar modelos de mucoviscidosis que reproducen algunos de los signos de la afección en humanos, y fue posible probar la viabilidad, la eficacia y la inocuidad de las primeras tentativas de terapia génica. Desde entonces, la técnica de recombinación homóloga se ha refinado considerablemente, y ahora es posible controlar la disfunción de un gen en el tiempo y en el espacio, como si estuviéramos accionando un interruptor. Volviendo al ejemplo de la mucoviscidosis, o del gen responsable, el crtf, ha sido posible estudiar las consecuencias de la ausencia de su expresión en tipos de células específicos. Como esta técnica se basa en la homología de la secuencia del fragmento introducido con la del gen mutado, uno de los requisitos para poder emplearla es conocer, al menos, una parte de la secuencia del adn del gen que se quiere modificar.
A pesar de estos avances, los modelos animales no siempre constituyen una buena copia fenotípica de las afecciones humanas. Por ejemplo, los ratones mdx, que al igual que los niños con miopatía de Duchenne están desprovistos de distrofina muscular, presentan un diagnóstico vital normal aun cuando sus músculos presentan ciertas lesiones causadas por el proceso necrótico característico de esta enfermedad. En algunos casos es posible contar con otros modelos animales, como ciertas afecciones articulares o de hipertensión, en las cuales las ratas desarrollan una semiología cercana a la enfermedad humana. Sin embargo, la transgénesis es aún una técnica difícil de realizar en especies que difieren del ratón, como ratas, conejos y vacas. Una de las aplicaciones de la transgénesis podría ser el uso de los órganos de animales transgénicos para realizar xenotransplantes. Aunque esta idea ya no sea ciencia ficción pura, la técnica aún presenta grandes dificultades de orden inmunológico y de seguridad que será necesario vencer antes de poder ponerla en práctica.
Comprender mejor para curar mejor
El hecho de haber identificado al gen responsable de una afección no implica que se comprenda su función o su participación en la fisiopatología, sin embargo, es necesario establecer los lazos que unen a la estructura del gen y a la proteína con los efectos deletéreos que produce su ausencia, para poder diseñar estrategias terapéuticas dirigidas y eficaces. Un ejemplo espectacular y reciente que ilustra este proceso es el de los trabajos emprendidos por dos equipos franceses en torno a una enfermedad muscular, la ataxia de Friedreich, un padecimiento que sufre una persona de cada cincuenta mil en Europa. El grupo de Jean Louis Mandel y Michel Koenig, en Estrasburgo, identificó el gen responsable de esta enfermedad y bautizó a la proteína para la cual codifica, pero cuya función se desconoce, frataxina. Un año más tarde, el equipo de Arnold Munich y Agnes Rôtig, en París, observó una deficiencia en las proteínas fierro-azufre mitocondriales en las biopsias de endiomiocardio de los pacientes con ataxia de Friedreich. Con estas dos claves era posible describir la secuencia fisiopatológica de la enfermedad aunque no se hubiese descubierto la función precisa de la frataxina: acumulación de fierro mitocondrial, pérdida de las proteínas fierro-azufre y sobreproducción de aniones superóxidos tóxicos para las células. Siguiendo esta lógica, se probaron in vitro numerosas sustancias farmacológicas que pudieran intervenir en este ciclo. La vitamina c, que aumenta la producción de fierro reducido, aumentó la toxicidad; el desferral desplazó el fierro responsable de la destrucción de las proteínas fierro-azufre solubles de las membranas, y un agente antioxidante, el idebenone, que no reduce el fierro, protegió a las enzimas, tanto a las solubles como a las membranales. El tratamiento con idebenone permitió mejorar espectacularmente la hipertrofia cardiaca de los tres primeros pacientes tratados, y actualmente, solamente tres años después del descubrimiento del gen, ya se está llevando a cabo un ensayo clínico con más de cincuenta enfermos.
Desgraciadamente esto no sucede así para todas las enfermedades. Aunque el gen de la distrofina muscular de Duchenne fue identificado desde hace más de diez años, no se ha podido diseñar una terapia eficaz para los pacientes con este padecimiento. Sin embargo, numerosos trabajos han permitido la consolidación del conocimiento acerca de las proteínas de esta familia. Un equipo inglés encontró una proteína análoga, la utrofina, que se expresa durante la vida fetal debajo de la membrana muscular al igual que la distrofina, pero que desaparece casi totalmente y sólo se expresa en las uniones neuromusculares después del nacimiento. Experimentos con modelos animales mostraron que la utrofina es capaz de reemplazar a la distrofina en el músculo adulto, reforzando así la idea de la existencia de una homología funcional entre ambas proteínas. Uno de los ejes de investigación terapéutica consiste en tratar de estimular la reexpresión de la utropina fetal con el fin de prevenir la necrosis muscular.
Las estrategias terapéuticas derivadas del conocimiento del genoma no se restringen a las enfermedades monogenéticas o cancerosas. También se pueden aplicar para tratar enfermedades inducidas por agentes infecciosos. ¿O no fue acaso el conocimiento del genoma del virus del sida lo que permitió imaginar el eficaz tratamiento con antiproteasas?
Una mina de medicamentos
Mucho antes de la revolución del genoma los genes ya servían como una fuente indiscutible de proteínas medicamentosas. Gracias a la ingeniería genética y al conocimiento de la secuencia protéica y de su maduración postraduccional —que en ocasiones es esencial para su funcionamiento— ha sido posible desarrollar terapias de sustitución, es decir, la administración de la proteína “sana” para compensar a la que falta o que está mutada. Para obtener una proteína determinada en grandes cantidades se introduce el gen en un organismo, desde una bacteria hasta un mamífero, para que su maquinaria de transcripción y de traducción la sintetice. Esta técnica de síntesis de proteínas es preferible a la de purificación a partir de tejidos animales o humanos que ocasionaron graves accidentes. Entre la larga lista de proteínas que se producen con este método, también llamadas proteínas recombinantes, se encuentran la insulina, la eritropoietina, el factor viii, el factor ix, las enzimas lisosomales, la hormona de crecimiento, las citoquinas y el interferón.
En la era de la automatización y de la informatización intensivas, la secuenciación completa de nuestro genoma puede significar la posibilidad de identificar todos los sitios de intervención terapéutica o las moléculas protéicas codificadas por esos genes. En efecto, los genes codifican para las enzimas, los receptores o los canales, que son las proteínas sobre las cuales actúan y actuarán los medicamentos de hoy y del mañana. Un importante esfuerzo industrial ha sido puesto en marcha para identificar moléculas naturales y sintéticas capaces de interaccionar y modular la actividad de estos sitios “blanco”. Se han emprendido varios programas para identificar nuevos moduladores químicos de la expresión génica, en particular, se busca aumentar la actividad específica de los promotores de genes que tengan un interés terapéutico. Este estudio se puede realizar en cultivos de células que contengan en su genoma la región promotora del gen que se quiere analizar acoplada a un gen marcador. De esta manera se pueden probar varios miles de moléculas en cadenas automatizadas, lo que se conoce como highthrouput screening o tamiz de alto rendimiento. La modulación de la expresión del gen marcador se analiza por medio de sistemas ópticos integrados a procesadores informáticos. Una vez que se identifica una molécula activa se estudia su biodisponibilidad, toxicidad y metabolismo. En nuestro ejemplo de la distrofia muscular de Duchenne, una vía terapéutica posible consistiría en buscar moléculas capaces de estimular la reexpresión de la utropina bajo la membrana muscular como en el estado fetal, así como se logró la reexpresión de la hemoglobina fetal por medio de butirato, hidroxiurea y eritropoietina en la drepanocitosis.
Así como la genómica se refiere a la genética en la época de los programas de los genomas, la farmacogenómica es una nueva forma de ver la farmacogenética a una gran escala. La idea subyacente es que el desarrollo de los métodos de estudio genéticos de los individuos permitirá desembocar en una terapéutica personalizada, adaptada a cada caso en función de la sensibilidad individual a los efectos terapéuticos e iatrogénicos de los medicamentos. La fabricación de chips de adn, que inmoviliza en un soporte sólido sondas, permite explorar miles de eventos genéticos en poco tiempo. Por ejemplo, se podrá detectar la presencia de mutaciones activadoras de oncogenes, alteraciones de antioncogenes, modificación de genes de susceptibilidad, etc.
El empleo de los chips nos permite esperar que las personas con determinadas afecciones podrán recibir un tratamiento mejor adaptado a la forma etiológica de su enfermedad y de su “acervo genético”, en términos de eficacia y de minimalización de riesgos de toxicidad.
La terapia génica: un concepto evolutivo
La terapia génica podría definirse de varias maneras, pero tal vez el significado menos restrictivo sea el de la utilización de un gen como molécula medicamentosa. En realidad no es un concepto nuevo, pues en un principio se veía la terapia génica como un transplante de genes al igual que uno de órganos, en donde el órgano sano suple la función del defectuoso, por lo que se pensaba que este tipo de terapia estaría reservado a las enfermedades genéticas. De hecho, además de las afecciones hereditarias, todos los tratamientos de sustitución que emplean proteínas recombinantes podrán ser reemplazados por la transferencia de los genes que codifican para esas proteínas. Además de esta terapia génica de “prótesis”, la perspectiva de poder llevar a cabo una reparación directa de los genes mutados ha dejado de ser ciencia ficción.
¿Terapia germinal o terapia somática?
La transgénesis, como se aplica para fines terapéuticos en modelos murinos (de ratón) de enfermedades humanas, constituye el arquetipo de lo que llamamos terapia germinal. El gen introducido desde una etapa precoz de la embriogénesis estará presente en todas las células del organismo y, por consiguiente, se transmitirá a la descendencia. En realidad hay muy pocas indicaciones en las que se podría aplicar al hombre, ya que solamente las afecciones dominantes en estado homocigoto, que son excepcionales, se verían beneficiadas por un tratamiento de este tipo. En todos los demás casos una proporción de la descendencia no son portadores de la mutación, y en consecuencia basta con transplantar al útero materno los embriones sanos, en vez de introducir un gen corrector cuyo efecto terapéutico en los embriones mutados es incierto.
En cambio, la terapia génica somática busca corregir una mutación o introducir un nuevo gen en un tejido determinado, o incluso en un tipo celular específico, sin modificar la herencia del paciente. Las aplicaciones para este tipo de terapia son muy variadas, y a partir de este momento sólo nos referiremos a este tipo de terapia génica a lo largo del texto.
¿Transferencia de células o de genes?
De manera un poco esquemática se puede decir que hay dos estrategias distintas para introducir un gen en un organismo: la transferencia directa o la transferencia de células genéticamente modificadas ex vivo, es decir, transformadas fuera del organismo. La estrategia directa, in vivo, parece ser técnicamente más sencilla puesto que consiste en aportar directamente el gen de interés al órgano afectado. Ésta se utilizará preferentemente en los casos en los que se disponga de una vía fácil de acceso al tejido blanco, cuando las células que se quieran tratar no se puedan extraer, o cuando éstas se encuentren diseminadas por todo el organismo. La mucoviscidosis, que afecta sobre todo a las células epiteliales del tracto traqueobronquial, ciertas enfermedades neurodegenerativas, así como la miopatía de Duchenne, son algunos ejemplos en los cuales se podría aplicar la terapia génica in vivo.
La estrategia celular, en cambio, consiste en aislar células del paciente, cultivarlas ex vivo e insertarles, generalmente por medio de vectores virales, el gen terapéutico. Esta estrategia se parece a los autotransplantes. Entre las enfermedades susceptibles a ser tratadas por este método y para las cuales los primeros ensayos clínicos, desgraciadamente aún raros, dieron resultados biológicos efectivos, se encuentran el déficit en adenosina desaminasa y la hipercolesterolemia familiar debida a la falta de receptores ldl. En el caso del déficit en adenosina desaminasa, los linfocitos aislados de niños afectados fueron infectados con un vector retroviral que contenía una versión normal del gen y, posteriormente, se volvieron a introducir en los pacientes. Las respuestas inmunológicas de los enfermos tratados mejoraron notablemente de manera durable. En el caso de la hipercolesterolemia, los hepatocitos de los pacientes se aislaron a partir de una biopsia hepática, se pusieron en cultivo y se indujo su proliferación al mismo tiempo que se infectaban con un retrovirus que contenía el gen del receptor ldl. Estos hepatocitos tratados fueron reinyectados a los pacientes por vía intraportal, sin embargo, el tratamiento no resultó muy eficaz. Posiblemente esto se debe a que la técnica, sin duda muy pesada, está además limitada por el número de células que se pueden corregir y reimplantar.
Cualquiera que sea la estrategia que se emplee, la directa o la celular, hay que notar que es posible hacer secretar una proteína potencialmente terapéutica a un tipo celular que normalmente no la produce. Esto permite escoger un tipo celular que sea capaz de secretar de manera eficaz una proteína terapéutica, aunque no sea el blanco principal de la enfermedad, lo cual resulta en más posibilidades terapéuticas. De la misma forma, las células modificadas pueden ser reimplantadas en un lugar que no corresponda con el tejido de origen. Por ejemplo, algunos experimentos con mamíferos grandes, como los perros, mostraron que es posible programar fibroblastos para que secreten una proteína lisosomal. Cuando se implanta en la cavidad peritoneal del perro un tejido sintético inerte constituido de fibras de colágeno y factores de crecimiento, llamado organoide, que espontáneamente se vasculariza, los fibroblastos secretan de manera activa la enzima.
Medicina regenerativa
Uno de los factores limitantes es la necesidad de obtener un buen número de células corregidas para poder generar un efecto terapéutico. Por lo tanto, si se les pudiera conferir alguna ventaja para su proliferación se incrementaría la eficacia de la estrategia. En este sentido, los resultados obtenidos por el equipo de Alain Fisher en el hospital Necker de París en el tratamiento de niños con un déficit inmunológico, son particularmente interesantes. En este caso, la introducción del gen normal en las células hematopoiéticas de los pacientes produjo una ventaja proliferativa a las células modificadas sobre las que residen en la médula. Esto permite explicar la extraordinaria eficacia que obtuvieron con el tratamiento a pesar de que el número inicial de células corregidas era muy bajo. Este concepto de “medicina regenerativa” no se limita al tejido hematopoiético, también se ha demostrado que funciona en el tejido hepático del ratón. Como el hígado posee la capacidad espontánea de regenerarse, es posible, confiriéndoles una ventaja selectiva a los hepatocidos reimplantados, inducir la proliferación de los hepatocitos corregidos en detrimento de los hepatocitos residentes y, de esta manera, asistir a la población progresiva del hígado con células modificadas. Este concepto tendrá, probablemente, grandes aplicaciones en la terapia génica del tercer milenio.
Desde la perspectiva del empleo de células con una ventaja proliferativa, las células madre son muy atractivas, ya que constituyen, al menos en teoría, un reservorio casi ilimitado de células diferenciadas potencialmente útiles en terapia génica. Desde algunos años parece que nuestro patrimonio de células madre de hígado, músculo e, incluso, de cerebro, es mucho más grande de lo que imaginábamos.
Así, hemos podido observar células neuronales que se diferencian en células hematopoiéticas, células hematopiéticas en hígado o músculo, e incluso hemos podido aislar células de músculo esquelético capaces de colonizar la médula. Este tipo de células parecen adaptar su comportamiento al medio en el que se encuentran, como si fueran una especie de camaleón. Ser capaces de aislar, cultivar y hacer proliferar este tipo de células sin duda permitiría revolucionar la terapia por autotransplante de células, genéticamente modificadas o no, de todos los tipos de afecciones para las cuales la única posibilidad terapéutica es el transplante.
Terapia génica aditiva o cirugía reparadora
Los primeros esfuerzos de terapia génica fueron dirigidos a las enfermedades genéticas. La adición de una copia normal del gen permite pasar, al menos en los casos de las enfermedades recesivas, de un estado homocigoto que presenta la enfermedad, a un estado heterocigoto fenotípicamente sano. Sin embargo, la necesidad de transmitir el gen a todas las células del tejido afectado por la enfermedad y de que la proteína terapéutica se exprese durante toda la vida del paciente ha restringido de manera considerable la eficacia terapéutica de los ensayos clínicos planeados hasta ahora. Si bien, el concepto es muy elegante, su realización es, por lo pronto, delicada. Otra posibilidad, considerada por mucho tiempo como mítica, se ha abierto recientemente: se trata de la posibilidad de reparar el sitio del gen mutado, como un tipo de cirugía estética. Esta posibilidad, realmente revolucionaria, de poder cambiar la porción mutada de un gen por su contraparte sana, se basa en el hecho de que las dobles hélices de arn-adn son más estables que las de adn-adn. En los casos en los que se conoce la secuencia mutada basta con introducir una secuencia muy corta de arn-adn en las células donde la mutación es deletérea, para corregir la zona mutada. Esta técnica, también conocida como quimeroplastia, tiene la gran ventaja de que no introduce ninguna secuencia exógena al genoma de las células tratadas. Hasta ahora solamente se ha probado en animales pero ya ha dado resultados espectaculares en distintos modelos de enfermedades humanas, como la hemoglobina, donde fue posible corregir 30% de las células hepáticas, o en un modelo de Crigler-Najjar, una hiperbilirrubinemia severa. Sin embargo, es necesario recalcar que esta cirugía sólo se aplica a casos de mutaciones puntuales, ya que no funciona más que para un número muy pequeño de pares de bases, y que en un gran número de enfermedades cada paciente necesitaría su propia terapia.
El geneticista que desde hace más de veinte años clona genes e identifica las mutaciones responsables de las enfermedades genéticas sin poder realizar su sueño de repararlas, no podría esperar mejor recompensa a sus esfuerzos que la obtención de resultados equivalentes en humanos.
Un resultado promisorio
Frecuentemente se le reprocha a las estrategias de terapia génica el hecho de ser extremadamente caras y difícilmente generalizables. A pesar de ello, esta intensa actividad de investigación ha llevado a una aplicación marginal del ámbito de la prevención más que de la terapéutica, que podría revelarse como extremadamente eficaz y poco costosa: la vacunación. En efecto, ahí donde la terapia génica fracasa por causa de la eficacia restringida de la transferencia de genes y por los límites temporales de la expresión, la vacunación por inyección de adn desnudo, que no requiere más que una pequeña cantidad y una expresión transitoria del antígeno, podría constituir una vía futura. La inyección intramuscular de adn desnudo que codifica para los antígenos ha permitido desencadenar una respuesta inmune de tipo celular y humoral. El uso extensivo de este método para la vacunación antiviral en países en vías de desarrollo sería suficiente para justificar todos los esfuerzos emprendidos hasta este momento.
|
|||
|
Helène Gilgenkrantz
Jacques Emmanuel Guidotti
Axel Kahn
Institute Cochin de Génétique Moléculaire,
inserm u129, París.
Traducción
Nina Hinke
|
|||
| ← Regresar al índice ⁄ artículo siguiente → |
 |
 |
||||
|
|
|||||
|
Las metapoblaciones en la naturaleza, ¿realidad o fantasía?
|
|||||
|
María Teresa Valverde Valdés
conoce más del autor
|
|||||
|
|
← Regresar al índice ⁄ artículo siguiente → | ||||
|
Cuando estudiamos la dinámica de una población ecológica (entendida ésta como un grupo de organismos de la misma especie que coexiste en un espacio común) generalmente nos centramos en el análisis de los parámetros demográficos básicos, a saber, las tasas de natalidad, de mortalidad y de migración. Sin embargo, es muy común que este último término sea ignorado (i.e., la migración) y que se parta de la base de que los únicos procesos que determinan los cambios numéricos de una población son los nacimientos y las muertes de los individuos. La teoría metapoblacional se encuentra fincada en el hecho de que la migración entre las poblaciones de una misma especie no siempre es despreciable, sino que por el contrario, con frecuencia es de gran importancia, pues determina la dinámica espacial de los conjuntos de poblaciones de una misma especie a través del tiempo. Esta idea nos lleva directamente a la definición del concepto de metapoblación: las diferentes poblaciones de una especie no se encuentran aisladas en la naturaleza, sino que mantienen cierto contacto entre ellas a través de la migración de individuos; a estos conjuntos o grupos de poblaciones de una misma especie, interconectados por la migración de individuos entre ellas, se les conoce como metapoblaciones.
Una de las ideas implícitas en el concepto metapoblacional es la suposición de que las poblaciones que forman una metapoblación (llamadas “poblaciones locales”) no son eternas, sino que pueden extinguirse por diversas causas. De la misma forma, algunos de los sitios o “parches” del ambiente susceptibles de ser colonizados por la especie pueden encontrarse vacíos y la migración de individuos hacia ellos puede dar lugar a la formación de nuevas poblaciones. De esta manera, una metapoblación puede concebirse como un sistema dinámico en el que constantemente ocurren la formación y la extinción de poblaciones locales. Así, de manera análoga al estudio de una población, cuya dinámica se explica con base en las tasas de natalidad y mortalidad, el estudio de la dinámica de una metapoblación se basa en el conocimiento de las tasas de colonización y extinción de poblaciones locales en un contexto regional, y en el análisis de las condiciones que permiten un equilibrio entre ambos procesos. Dado que las especies que forman metapoblaciones se encuentran sujetas a altos riesgos de extinción de sus poblaciones locales, su permanencia a nivel regional depende del balance entre colonizaciones y extinciones. Surge el concepto metapoblacional Al analizar la historia del concepto metapoblacional, aparentemente nuevo en ecología, causa sorpresa encontrarse con el hecho de que dicho concepto data de finales de la década de los sesentas. De hecho, la idea básica de lo que en la actualidad se conoce como una metapoblación se encuentra ya en textos ecológicos clásicos, como el de Andrewartha y Birch, incluso anterior a esas fechas. Sin embargo, no es sino hasta finales de los sesentas que Richard Levins, un entomólogo interesado en el control de plagas, “bautiza” a las metapoblaciones y formaliza el concepto mediante, de la propo sición del primer modelo metapoblacional, que posteriormente daría pie al desarrollo de lo que ahora conocemos como teoría metapoblacional. Es interesante analizar el contexto histórico en el que Levins propone, por vez primera, la existencia de metapoblaciones en la naturaleza: desde los años treintas y hasta finales de la década de los cincuentas se dio una controversia importante y de consecuencias muy fructíferas entre los ecólogos poblacionales, que tenía que ver con la discusión sobre los factores que regulan el tamaño de las poblaciones en la naturaleza. En esa época existían dos escuelas principales de pensamiento: a) una que sostenía que los factores reguladores del tamaño poblacional eran de naturaleza biótica (depredación, parasitismo, competencia) y que por lo tanto se manifestaban con mayor intensidad en poblaciones con densidades altas, y b) otra que planteaba que eran principalmente los factores abióticos (tormentas, perturbaciones, heladas, etcétera) y, por lo tanto, independientes de la densidad poblacional, los que mantenían a las poblaciones alrededor de ciertos valores de densidad. Una tercera alternativa que recibió poca atención en ese momento planteaba que los mecanismos de regulación poblacional no son tan eficientes como se suponía, y que si se seguía a una población por un tiempo suficientemente prolongado se observaría que, a la larga, tendería a la extinción. Entonces, la permanencia a nivel regional de especies con este tipo de comportamiento numérico debía estar determinada por un balance entre la extinción de poblaciones locales y la formación de poblaciones nuevas (a través de la migración) al interior de un “ensamble” de poblaciones locales efímeras.
No es coincidencia que haya sido precisamente Levins, un entomólogo especializado en el control de plagas, el que desarrolló esta idea y la plasmó en el modelo metapoblacional básico (que se presenta más adelante), pues si analizamos el desarrollo regional de cualquier plaga nos daremos cuenta de que ésta se comporta como metapoblaciones: las plagas no tienen mecanismos eficientes de control poblacional, sino que llegan a un lugar, consumen todos los recursos disponibles y posteriormente desaparecen. El hecho de que a pesar de estas extinciones muchas plagas sean tan difíciles de erradicar es un reflejo de que su permanencia está dada por un balance entre las extinciones y la formación de nuevas poblaciones a través de la colonización de sitios suceptibles de ser explotados. Algunos de los grandes aciertos de Levins al proponer el concepto metapoblacional son, por un lado, la descripción de esta dinámica mediante del planteamiento de un modelo matemático sencillo y, por otro lado, la sugerencia implícita de que posiblemente muchas otras especies, además de las plagas, presenten este tipo de dinámicas espacio-temporales, detectables únicamente por encima del nivel de análisis estrictamente poblacional. La propuesta de Levins El modelo de Levins considera especies que muestran patrones de distribución espaciales a manera de “parches” discretos y bien definidos en los que la especie puede vivir inmersa en una “matriz” de hábitat formada por ambientes no colonizables. El modelo considera únicamente los parches en los que la especie puede establecerse (por ejemplo, los árboles hospederos de una planta epífita en un bosque, o las charcas de agua que se forman en las praderas y que son colonizadas por ciertas especies de ranas), y describe el estado de la metapoblación en un momento dado en términos de la proporción de estos parches que se encuentran realmente ocupados por la especie y la proporción de parches que se encuentran vacantes. El modelo estima el cambio en la proporción de parches ocupados a través del tiempo, tomando en cuenta la tasa de colonización de parches (formación de poblaciones locales nuevas) y la tasa a la que las poblaciones locales se extinguen (ver recuadro).
Es importante hacer notar que el modelo metapoblacional de Levins es prácticamente idéntico en su forma a algunos de los modelos desarrollados en epidemiología para describir la dinámica de las infecciones por parásitos. Esto es reflejo de que, en ambos casos, se concibe a una especie (el parásito, en el caso de los modelos epidemiológicos) que va “infectando” ciertos ambientes colonizables mientras es erradicada de otros, generando la dinámica de colonización–extinción que caracteriza a ambos sistemas.
El modelo metapoblacional de Levins está basado en varios supuestos fundamentales: 1) la extinción sucede y, por lo tanto, la permanencia de una especie en una región debe depender de la colonización; 2) todos los parches disponibles tienen la misma probabilidad de ser colonizados, es decir, no hay un efecto significativo de la distribución espacial o del nivel de aislamiento de los parches; 3) la tasa de colonización es proporcional a la fracción de parches ocupados; 4) todas las poblaciones tienen la misma probabilidad de extinguirse, pues no hay un efecto del tamaño, la longevidad o la dinámica interna de las poblaciones locales, y 5) la probabilidad de extinción de cada población es independiente, ya que no ocurren extinciones correlacionadas.
Es claro que estos supuestos no se cumplen en las metapoblaciones que encontramos en la naturaleza, pero también es cierto que la mayoría de ellos pueden relajarse sin necesidad de hacer modificaciones conceptuales significativas al modelo. En realidad, utilizar un modelo metapoblacional con estas características no es muy distinto de la forma en la que se utiliza el concepto de “población” en biología poblacional: ninguna población real satisface cabalmente los criterios de ser una entidad panmíctica (en donde la reproducción puede darse entre todos los individuos que la conforman) y cerrada (sin migración), y a pesar de todo el concepto poblacional es útil e ilustrativo para abordar el estudio de una parte importante de la naturaleza viva. De la misma forma, el concepto de metapoblación ofrece una concepción particular sobre la estructura espacial de las especies en la naturaleza, que nos habla de la existencia de poblaciones locales discretas, conectadas a través de la migración de individuos entre ellas. Esta concepción, en sí misma, es de gran valor, pues considera una escala espacial (regional) que no había sido abordada por la ecología poblacional; enfatiza procesos que se dan en una escala temporal y espacial que no siempre es obvia para el ojo humano, y, además, constituye un salto desde la ecología descriptiva, que señala la existencia de patrones en la naturaleza, hasta la ecología analítica, que se preocupa por describir los procesos que dan lugar a dichos patrones. Extinción y tipos de metapoblaciones El concepto metapoblacional y, consecuentemente, toda la teoría que se desprende de él, dependen de la idea de que la extinción de las poblaciones locales ocurre (y de ahí la importancia de la migración para permitir la recolonización). En efecto, en la literatura se encuentran reportados un gran número de casos en los que se ha documentado la extinción de poblaciones locales de diversas especies de animales y plantas, lo cual sugiere que las dinámicas de colonización–extinción podrían ser importantes en algunos sistemas naturales. Las causas que han provocado estas extinciones pueden ser de tipo estocástico (es decir, debidas al azar) o de naturaleza determinística. Las extinciones estocásticas tienen que ver con tres tipos fundamentales de procesos azarosos: 1) aquellos que ocurren a nivel ambiental (tales como las variaciones en las condiciones climáticas, las heladas severas, las tormentas, los deslaves, los ciclones, los incendios, etcétera); 2) los que toman lugar a nivel de la dinámica interna de las poblaciones locales (i.e., “estocasticidad demográfica”, dada por variaciones aleatorias en el orden y magnitud de los nacimientos y muertes de individuos al interior de una población), y 3) los que tienen que ver con las características genéticas de las poblaciones (i.e., “estocasticidad genética”, en donde los procesos de deriva génica pueden provocar pérdida de variabilidad genética y depresión endogámica y llevar a una población a la extinción).
Las extinciones determinísticas (es decir, ocasionadas por procesos no azarosos) también parecen ser relativamente frecuentes. Éstas ocurren como producto de cambios direccionales en las condiciones del medio que poco a poco provocan que el parche ocupado por una especie se transforme en un sitio inhabitable. Esto ocurre, por ejemplo, en los hábitats de carácter efímero o temporal, tales como una fruta que cae al suelo en un bosque y se convierte en un parche susceptible de ser colonizado por larvas de insectos, hongos, etcétera, pero que a la larga desaparecerá. Otro tipo de extinciones determinísticas son las que se dan en especies que ocupan hábitats sucesionales tempranos dentro de algunas comunidades. Por ejemplo, muchas especies de plantas colonizan los claros que se forman en los bosques cuando ocurre una pequeña perturbación, como la caída de un árbol o una rama grande; estos claros se convierten en parches que pueden ser colonizados por especies de plantas que requieren de este tipo de condiciones ambientales para establecerse, sin embargo, al irse cerrando el claro a causa del crecimiento de la vegetación adyacente (proceso conocido como sucesión secundaria), el parche se convierte poco a poco en un sitio inhabitable y la población local que lo haya ocupado estará destinada a la extinción. Basada en la relevancia a nivel regional de los procesos de colonización–extinción para diferentes especies, Harrison propone la existencia de cuatro tipos de metapoblaciones: a) Metapoblaciones tipo isla-continente. Son aquellas en las que existe una población grande que funciona como la fuente principal de colonizadores de los parches pequeños cercanos a ella. En este tipo de metapoblaciones las extinciones se dan únicamente en las poblaciones pequeñas (las “islas”), mientras que la población grande (el “continente”) es prácticamente inmune a la extinción. Consecuentemente, sí existe una dinámica de colonización–extinción, pero ésta no determina la permanencia de la especie a nivel regional. Este tipo de metapoblaciones se han observado en algunas especies de mariposas. b) Metapoblaciones tipo mosaico. Son aquellas en las que la especie presenta, en efecto, una distribución a manera de parches discretos, pero entre los cuales la migración de individuos es tan generalizada que no se puede considerar a cada parche como una población local relativamente independiente. El conjunto de parches forma, en realidad, una entidad demográfica única e integrada y el alcance tan generalizado de la migración no permite la extinción. Algunas especies sucesionales, como las que se describieron en el párrafo anterior, parecen tener este tipo de estructura metapoblacional. c) Metapoblaciones no equilibradas. Muchas especies que originalmente no presentaban una estructura metapoblacional se han visto recluidas, en las últimas décadas, a pequeños fragmentos de hábitat natural rodeados de ambientes alterados, como los campos de cultivo o los pastizales de uso ganadero. Estas especies presentan ahora muchas de las características propias de las metapoblaciones, tales como la existencia de parches colonizables rodeados de una matriz de hábitat no colonizable y la alta ocurrencia de extinciones locales. Sin embargo, la re-colonización de los parches vacantes se ve limitada por la baja capacidad de migración de los individuos y, por lo tanto, la tasa de extinción excede a la tasa de colonización. Así, este tipo de metapoblaciones se presentan en el contexto de una disminución generalizada en la abundancia de muchas especies, determinada por el deterioro paulatino del hábitat. Muchas especies de mamíferos grandes, comoalgunos felinos y osos, presentan este tipo de metapoblaciones. d) Metapoblaciones clásicas. Son aquellas que se acercan en mayor medida a la idea de metapoblación propuesta en el modelo original de Levins. En estos sistemas existe un balance real entre colonizaciones y extinciones, que es lo que determina la permanencia de la especie a nivel regional. Se han documentado muy pocos casos que cumplan estrictamente con estas características; uno de los más notables corresponde a un sistema de charcas que son colonizadas por una especie de rana y en donde son frecuentes tanto la extinción de poblaciones locales como la formación de poblaciones nuevas. Metapoblaciones y conservación El concepto metapoblacional propuesto por Levins no tuvo mucho eco en la comunidad de ecólogos en su momento, sino que pasó prácticamente inadvertido hasta ya bien entrada la década de los ochentas, cuando se retomó y se comenzó a elaborar poco a poco para dar paso a la actual teoría metapoblacional. Los temas de interés entre los ecólogos poblacionales de los sesentas eran hasta cierto punto distintos de los que ahora constituyen el foco de atención de los ecólogos contemporáneos, y la visión de Levins no adquirió la generalidad necesaria como para llamar la atención en un círculo amplio de investigadores. En contraste, uno de los temas de vanguardia de la ecología contemporánea es precisamente la dinámica y la estructura espacial de los sistemas ecológicos. Es quizá por esta razón que la teoría metapoblacional parece haber encontrado su nicho natural en la ecología moderna, al ofrecer una visión particular de la dinámica espacial entre grupos de organismos de la misma especie. La teoría metapoblacional actual va incluso más allá del nivel exclusivamente uni–específico y aborda el área de las interacciones ecológicas, como la competencia y la depredación, planteando la forma en la que ciertos sistemas de pares de especies (competidoras y depredador–presa) pueden coexistir en equilibrio gracias a la existencia de estructuras metapoblacionales. Sin embargo, más que en el ámbito de la ecología tradicional, la teoría metapoblacional ha tenido un impacto importante en el área de biología de la conservación. Quizá el auge de esta disciplina en tiempos recientes es otra de las razones que han determinado el resurgimiento de la teoría metapoblacional. Un hecho que bien da cuenta de esta asociación entre la biología de la conservación y la teoría metapoblacional es que esta última aborda formalmente el tema de la extinción; además, al analizar la dinámica espacial de especies de interés particular, ofrece una serie de criterios ecológicos sólidos para apoyar el diseño de reservas, corredores ecológicos, áreas protegidas y demás. Respecto de este último punto, es interesante notar que durante la década de los años setenta el paradigma de la biología de la conservación, sobre todo en lo referente al diseño de reservas, fue la teoría de biogeografía de islas, planteada por MacArthur y Wilson en 1967, mientras que ahora es la teoría metapoblacional la que generalmente se invoca para ofrecer criterios ecológicos en la toma de decisiones sobre áreas protegidas. Para entender las causas que han determinado este cambio de paradigmas es necesario analizar la contribución particular de cada una de estas dos teorías a la biología de la conservación.
Uno de los aspectos más importantes de la teoría de biogeografía de islas que posteriormente fue retomado por la biología de la conservación, fue la idea de que existe una relación entre el área ocupada por una cierta comunidad ecológica y el número de especies que puede mantener. Además, MacArthur y Wilson analizaron la relación entre la distancia a la que una “isla” (un fragmento aislado de hábitat colonizable) se encuentra del “continente” (es decir, de la fuente de colonizadores), y la tasa de llegada y desaparición de especies de esa isla, sugiriendo que el número de especies presentes en ella llegaría a un cierto equilibrio, dado precisamente por el área de la isla. Estas ideas parecían enfatizar el uso de determinadas estrategias, relacionadas con el tamaño, forma y disposición de las áreas protegidas, destinadas a maximizar el número de especies que albergarían. A partir de esto, los conservacionistas comenzaron a manejar una serie de “reglas” para el diseño de reservas que, incluso, a la fecha, se asocian con la teoría de biogeografía de islas. Algunas de estas “reglas” son, por ejemplo, la noción de que un área grande puede albergar una mayor diversidad que un área pequeña, o que áreas contiguas conectadas por corredores presentan más especies que áreas aisladas de tamaño equivalente. Nótese que a pesar de que existe una cierta similitud en el contexto en que se plantean las teorías metapoblacional y de biogeografía de islas (pues ambas visualizan el ambiente como un conjunto de parches colonizables rodeados de una matriz de ambiente no colonizable, además de que enfatizan el análisis de las tasas de colonización y extinción), éstas abordan aspectos muy diferentes de la ecología. La teoría de biogeografía de islas tiene que ver con la dinámica de conjuntos de especies y con el análisis de los factores que determinan la riqueza específica al interior de los parches colonizables presentes en una cierta área. La teoría metapoblacional, por su parte, analiza la dinámica espacial de una especie en particular y estudia las condiciones que afectan los cambios en el número de parches ocupados por dicha especie. Dicho en otras palabras y de manera un tanto simplista, la teoría metapoblacional forma parte de la ecología de poblaciones, mientras que la teoría de biogeografía de islas es claramente parte del ámbito de la ecología de comunidades. En resumen, estas dos teorías buscan respuestas a preguntas esencialmente diferentes y, por lo tanto, su contribución a la biología de la conservación, sobre todo en lo relacionado con el diseño de reservas, no es ni siquiera comparable, pues abordan aspectos distintos de la ecología. La teoría metapoblacional ofrece elementos para establecer criterios útiles para la creación de reservas, pero no desde la óptica de la diversidad de la comunidad en su conjunto, sino de la permanencia de especies particulares a nivel regional a través del conocimiento de la dinámica espacial de sus poblaciones. El cambio de paradigmas al que se ha hecho referencia con anterioridad parece tener varias causas. Una de ellas está relacionada con el hecho de que, como era de esperarse, no se han encontrado suficientes evidencias empíricas que apoyen las predicciones derivadas de la teoría de biogeografía de islas. Por otro lado, la biología de la conservación parece estar dando un giro del enfoque comunitario al poblacional en los últimos años (en el cual puede incluirse el reciente interés por los aspectos genéticos de las poblaciones), lo cual ha favorecido el auge de la teoría metapoblacional. Además, la adquisición de un enfoque metapoblacional ha permitido analizar los paisajes naturales como “redes” o sistemas de reservas, y no tanto como reservas aisladas, haciendo énfasis precisamente en las dinámicas espaciales de los sistemas naturales, que constituyen una de las preocupaciones fundamentales de la ecología moderna. ¿Por qué este enfoque? La breve exposición hecha en los párrafos anteriores sobre la importancia de la teoría metapoblacional en ecología de la conservación no es exhaustiva, ni pretende serlo; sin embargo, sí señala algunos de los aspectos de mayor relevancia que permiten explicar su utilidad desde el punto de vista teórico y conceptual. Ahora, es necesario tomar en cuenta que las metapoblaciones, como entidades reales en la naturaleza, pueden ser muy difíciles de delimitar, estudiar, o sencillamente observar, pues no nada más se presentan a escalas espaciales generalmente difíciles de manejar, sino que, además, prácticamente nunca cumplen con los supuestos del modelo metapoblacional original. Por otro lado, el movimiento de individuos entre poblaciones de una misma especie puede ser extremadamente difícil de cuantificar y describir, lo cual es una limitación muy seria si se toma en cuenta que la migración es una de las condiciones básicas del concepto metapoblacional. Todo esto ha llevado a ciertos autores, como Susan Harrison, a cuestionar la utilidad del concepto en su forma original un tanto rígida, a causa de la falta de evidencia empírica que lo apoye. De ahí el título del presente artículo, que hace referencia precisamente a esta crítica. Para responder en alguna medida a la crítica de Harrison, me gustaría retomar la relación entre los conceptos teóricos de población y de metapoblación, con relación a que ninguno de ellos refleja realmente ningún sistema natural en el sentido estricto de la palabra y que, a pesar de todo, ambos nos han sido útiles para describir y estudiar una parte importante de la naturaleza viva. Esta analogía, además, enfatiza uno de los puntos que me parece importante comentar en el presente artículo: el hecho de que los conceptos ecológicos no tienen que reflejar la realidad al cien por ciento para sernos útiles o para que los escojamos como una representación válida de algún aspecto de la realidad. Muchos de estos conceptos representan modelos de la realidad y, como tales, no la reflejan (ni pretenden hacerlo) en toda su magnitud y complejidad, sino que constituyen versiones simplificadas de los sistemas, fenómenos o procesos que nos interesa abordar; el único objetivo de esta simplificación es ofrecer una mayor accesibilidad al objeto de estudio en cuestión. En el caso del concepto metapoblacional, lo que es importante rescatar es esta concepción de que los grupos de poblaciones locales discretas de una especie, conectados a través de la migración de individuos entre ellas, pueden ser analizados a una escala espacial regional, por encima de la escala poblacional. Quizá no sea tan importante determinar con exactitud cuánta migración hay entre las poblaciones de algún sistema que nos interese estudiar, ni qué alcance espacial tiene esta migración, sino más bien decidir si al utilizar una visión metapoblacional vamos a alcanzar una mejor comprensión de ciertos procesos ecológicos que pasaríamos por alto si nos limitaramos a una visión exclusivamente poblacional. Desde un punto de vista poblacional o demográfico, nos podríamos formular la siguiente pregunta: ¿es válida la suposición de que las poblaciones son sistemas cerrados en donde el efecto de la migración puede ignorarse? Siempre que la respuesta a esta pregunta sea no, entonces podemos estar seguros de que el uso de una visión metapoblacional enriquecerá sustancialmente nuestra comprensión del sistema bajo estudio. |
|||||
|
Referencias bibliográficas
Andrewartha, H. y L. Birch. 1954. The distribution anda abundance of animals., Chicago, University of Chicago Press.
Hanski, I. y D. Simberloff. 1997. “The metapopulation approach, its history, conceptual domain, and application to conservation”, en Hanski, I. y M. E. Gilpin, eds., Metapopulation biology. Nueva York, Academic Press Inc., pp. 5-26. Harrison, S. 1991. Local extinction in a metapopulation context: an empirical evaluation. Biological Journal of the Linnean Society, 42 pp. 73-88. Levins, R. 1969. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bulletin of the Entomological Society of America, 15, pp. 237-240. Levins, R. 1970. Extinction. Lectures on Mathematics in the Life Sciences, 2, pp. 75-107. MacArthur, R y E. Wilson. 1967. The theory of island biogeography., Princeton, Princeton University Press. |
|||||
|
María Teresa Valverde Valdés
Facultad de Ciencias,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo →
Valverde Valdés, Maria Teresa. (1999). Las metapoblaciones en la naturaleza, ¿realidad o fantasía? Ciencias 53, enero-marzo, 56-63. [En línea] |
|||||
| ← Regresar al índice ⁄ artículo siguiente → |
 |
 |
|||
|
|
||||
|
Manuel Sandoval Vallarta y la física en México
|
||||
| Alfonso Mondragón
conoce más del autor
|
||||
|
|
← Regresar al índice ⁄ artículo siguiente → | |||
|
En el otoño de 1932, Manuel Sandoval Vallarta, un físico que impartía cátedra en el Instituto Tecnológico de Massachusetts (mit), propuso en Chicago la idea de un experimento decisivo para determinar la naturaleza de la radiación cósmica. Unas semanas después, en la ciudad de México, Luis W. Álvarez llevó a cabo el experimento y descubrió que la radiación cósmica primaria está constituida principalmente por protones y núcleos atómicos. En los años siguientes, un grupo dirigido por Sandoval Vallarta y por Georges Lemaître formuló y desarrolló la teoría de los efectos geomagnéticos en los rayos cósmicos, lo que le valió un amplio reconocimiento mundial. Años después, Manuel Sandoval Vallarta, junto con sus colaboradores, fue nominado al premio Nobel.
A pesar de vivir en el extranjero, Sandoval Vallarta ejerció directa e indirectamente una fuerte influencia en la organización y desarrollo de la física mexicana. En colaboración con Alfredo Baños, su alumno en el mit, organizó en 1937 el primer grupo de mexicanos dedicado a la investigación de la física moderna. De este modo, la fundación de la Facultad de Ciencias, en 1938, y de los Institutos de Física, en 1939, y de Geofísica, una década después, quedó indisolublemente asociada a la investigación teórica y experimental de la radiación cósmica con la que se inició el desarrollo contemporáneo de la física en México.
Manuel Sandoval Vallarta nació el 11 de febrero de 1899 en la ciudad de México. Su padre, Pedro Sandoval Gual, licenciado en derecho, fue director de la Lotería Nacional. Su madre, Isabel Vallarta Lyon, era hija del distinguido jurista y político liberal Ignacio L. Vallarta. Sandoval Vallarta pasó su infancia y adolescencia en el seno de esa antigua y distinguida familia de la alta burguesía mexicana, en la que se entremezclaban las tradiciones conservadoras y liberales con el amor a la ciencia y a la cultura. En 1912 ingresó a la Escuela Nacional Preparatoria, que en aquellos años era uno de los baluartes de la educación positivista en México. El profesor de física era don Juan Mancilla y Ríos, y el de cosmografía don José de las Fuentes. Estos dos excelentes profesores despertaron el interés del joven Sandoval Vallarta por los estudios de la física. Fue, sin embargo, el profesor de matemáticas, don Sotero Prieto, quien influyó de manera decisiva para definir la vocación de Sandoval Vallarta. El genuino interés que Sotero Prieto tenía por las matemáticas y por la física teórica lo llevaron a organizar seminarios y grupos de estudio con sus alumnos. Ahí se presentaban y discutían algunas de las ideas más novedosas y avanzadas de la física teórica de su tiempo, que por su nivel quedaban fuera del programa del curso regular.
A los dieciséis años Sandoval Vallarta decidió ir a estudiar con sir Joseph Larmor en la Universidad de Cambridge, Inglaterra. Sin embargo, no pudo hacer el viaje porque entre 1916 y 1917 la Primera Guerra Mundial estaba en su apogeo y la travesía por el Atlántico era una aventura muy arriesgada. Después de consultar con varios amigos, su padre le aconsejó que fuera a estudiar a la Universidad de Harvard o al Instituto Tecnológico de Massachusetts, ambos ubicados en la ciudad de Boston. Así, en agosto de 1917, realizó los exámenes de admisión en el Instituto Tecnológico, donde, después de aprobar el plan de estudios de la carrera de Ingeniería Eléctrica, obtuvo el grado equivalente a la licenciatura en 1921. Posteriormente se inscribió en el Doctorado de Ciencias en la especialidad de Física Matemática, obteniendo el grado de doctor en 1924, con una tesis titulada El modelo atómico de Bohr desde el punto de vista de la relatividad general y el cálculo de perturbaciones, que fue escrita bajo la supervisión del profesor H. B. Wilson.
En 1923, mientras era candidato a obtener el grado de doctor, fue nombrado ayudante del profesor Vannevar Bush. En ese puesto, inicialmente hizo un estudio experimental de la transmisión de corrientes de alta frecuencia en conductores, pero muy pronto dirigió su atención a los problemas teóricos. En conexión con la teoría necesaria para explicar los experimentos, Bush, desde 1921, le había sugerido que investigara el fundamento matemático del cálculo operacional de Heaviside, que aprendían los ingenieros como una receta práctica para encontrar la solución de las ecuaciones diferenciales asociadas a los circuitos eléctricos. Sandoval Vallarta demostró, en forma matemáticamente rigurosa, que el método era correcto y lo utilizó para calcular el estado transitorio de una línea de transmisión con circuitos terminales; un problema que, al igual que sus investigaciones experimentales, estaba dirigido a mejorar la transmisión de mensajes telegráficos y telefónicos por un cable submarino. Este método también aplicó al estudio de las oscilaciones mecánicas de sistemas complejos. Simultáneamente desarrollaba su ambicioso proyecto de investigación en física teórica, cuyo tema general abarcaba los problemas más importantes de esta ciencia en esa época: la Teoría General de la Relatividad de Einstein y el estudio de la estructura atómica y la interacción de la materia con la radiación electromagnética. Los resultados de esos trabajos fueron publicados en 1925 y 1926 en el Journal of Physics and Mathematics y The Physical Review. La calidad de sus trabajos, publicados antes de que cumpliera veintisiete años de edad, le dieron fama como uno de los investigadores jóvenes más brillantes, rigurosos y objetivos de su tiempo.
En 1927, Manuel Sandoval Vallarta ganó una beca de la fundación Guggenheim, situación que le permitió ir a la Universidad de Berlín, centro mundial de la física en esa época. Los profesores de física eran Albert Einstein, Max Plank, Erwin Schrödinger y Max von Laue. Con Einstein tomó el curso de relatividad y con Plank el de teoría electromagnética. Schrödinger enseñaba la mecánica cuántica, descubierta apenas el año anterior. Además tomó el curso de epistemología que impartía Hans Reichenbach y, como estaba interesado en cuestiones de exégesis, también asistió al curso de von Harnak. En esos años conoció a Johan von Neumann, a Eugen Wigner y a varios de los jóvenes europeos que más tarde llegarían a estar entre los más eminentes físicos teóricos del mundo. En 1928, al terminar los cursos en Berlín, con ayuda de la misma beca fue a Leipzig, en donde estuvo un año. Ahí pudo conocer a Werner Heisenberg, descubridor de la mecánica cuántica en su forma matricial. Heisenberg, que entonces tenía veintisiete años, había sido nombrado profesor de física en ese año. Por su parte, Peter Debye era profesor de teoría molecular. En 1929, Manuel Sandoval Vallarta regresó al Instituto Tecnológico de Massachusetts y fue nombrado profesor ayudante.
El conocimiento profundo de la física atómica y sus técnicas matemáticas le permitieron asimilar fácilmente las nuevas ideas de la mecánica cuántica descubierta apenas entre 1925 y 1926 por Heisenberg y Schrödinger. La serie de trabajos sobre las Condiciones de validez de la macromecánica, La teoría de la parte continua del espectro de rayos X y La teoría relativista de la mecánica ondulatoria, realizados antes del viaje a Alemania, fue seguida de nuevos esfuerzos por encontrar una formulación de la mecánica cuántica compatible con la relatividad especial, llevados a cabo en colaboración con D. J. Struik y Nathan Rosen. Ese trabajo sirvió a Rosen para obtener la Maestría en Ciencias en el Instituto Tecnológico de Massachusetts bajo la dirección de Sandoval Vallarta. Entre 1921 y 1932, este último publicó una serie de artículos dedicados al estudio de la teoría unificada del electromagnetismo y la gravitación propuesta por Einstein en 1918. Manuel Sandoval Vallarta admiraba a Einstein y conocía a fondo la teoría de la relatividad general propuesta en 1916 por ese autor. Sin embargo, fue un severo crítico de los esfuerzos de Einstein en la dirección de un campo unificado. En colaboración con Norbert Wiener hizo notar que era más urgente unificar la relatividad especial y la mecánica cuántica que buscar una teoría unificada de la gravitación y el electromagnetismo. El tiempo les dio la razón, aunque hay que señalar que este último problema sigue siendo una de las metas no alcanzadas de la física teórica. Con el mismo Wiener, y posteriormente él sólo, demostró que el límite estático de las teorías de Einstein de 1928 y 1929 permitían únicamente soluciones triviales y no llevaban a la importante solución de Schwarzschild de las ecuaciones de la relatividad general.
Esos resultados tuvieron una influencia profunda en los trabajos posteriores de Einstein, quien no tardó en encontrar una nueva forma de la teoría, libre de los defectos señalados. Con Nathan Rosen, Sandoval Vallarta analizó la nueva versión de la teoría y encontró que tampoco era satisfactoria. La publicación de ese trabajo sirvió para estrechar la relación personal de Manuel Sandoval Vallarta con Einstein, que ya era cordial y amistosa. Nathan Rosen fue enviado al Instituto de Estudios Avanzados de Princeton a colaborar con Einstein, y de esa colaboración resultó, entre otros frutos, el famoso trabajo de Einstein, Podolsky y Rosen que critica a la mecánica cuántica.
En el verano de 1932, Don Manuel se encontraba en la ciudad de México cuando recibió la visita de Arthur H. Compton, quien venía de un largo viaje por el Pacífico, llevado a cabo con el propósito de medir la intensidad de la radiación cósmica en muchos lugares de diferente latitud. Había salido de California a Hawaii, y de ahí a Nueva Zelanda. Después tocó las costas de Perú y Panamá hasta llegar a México. En su viaje observó que la intensidad de la radiación cósmica cambia con la latitud geomagnética, siendo mínima cerca del ecuador geomagnético. Acompañado por Sandoval Vallarta, Compton realizó nuevas medidas en nuestro país: en Orizaba, Veracruz; en la ciudad de México, y, finalmente, en el Nevado de Toluca. De ese modo, don Manuel se enteró del descubrimiento de Compton: la observación del efecto de latitud indicaba que la radiación cósmica está constituida por partículas cargadas de electricidad que llegan a la Tierra desde el espacio exterior moviéndose a gran velocidad. Don Manuel mostró un gran interés en los resultados de Compton, pues en éstos vio la posibilidad de encontrar la confirmación observacional de las ideas sobre el origen y la expansión del Universo formuladas por su amigo y condiscípulo Georges Lemaître un año antes. Lemaître había descubierto que las ecuaciones de la relatividad general de Einstein tienen una solución en la que toda la masa y toda la energía del Universo están inicialmente concentradas en una región muy pequeña del espacio. De acuerdo con esto, propuso la hipótesis del átomo primigenio, según la cual la explosión de éste había dado origen al Universo en expansión que observan los astrónomos. La gran explosión inicial estaría acompañada de la emisión de fotones y partículas cargadas y neutras, cuya energía decrecería gradualmente con la expansión del Universo. Si se pudiera calcular el efecto del campo magnético terrestre, los rayos cósmicos darían información que podría comprobar la teoría de Lemaître.
Al final del verano de 1932, Sandoval Vallarta regresó a Massachusetts y de inmediato se puso a trabajar con Lemaître en el desarrollo de una teoría cuantitativa del movimiento de una partícula cargada de electricidad en el campo magnético terrestre. En noviembre de 1932, la teoría ya estaba lo suficientemente desarrollada como para conducir a un resultado importante: las partículas cargadas de electricidad que, moviéndose a gran velocidad, se acercan a la Tierra desde el espacio exterior, son desviadas de la dirección original de su movimiento por el campo magnético terrestre, de tal manera que llegan al observador como si entraran por un cono o embudo que se abre hacia un lado y no hacia arriba. Si las partículas tuvieran carga positiva, el cono se abriría hacia el oeste, en tanto que si las partículas tuvieran carga negativa el cono se abriría hacia el este.
En una conferencia dictada en la Universidad de Chicago en noviembre de 1932, Manuel Sandoval Vallarta propuso que se midiera el efecto de asimetría este-oeste. También sugirió que la observación se hiciera en la ciudad de México por su latitud magnética baja y por su gran altura sobre el nivel del mar. Arthur H. Compton recogió la sugerencia y envió a uno de sus discípulos, Luis W. Álvarez, quien años más tarde ganaría el premio Nobel por sus trabajos en otro campo, a que se encargara del experimento. En la azotea del Hotel Géneve, de la ciudad de México, Álvarez armó los contadores Geiger en la tapa de una caja de madera colocada en una carretilla de albañil; fijó la posición de la tapa de tal modo que los contadores apuntaran a un cierto valor del ángulo cenital e hizo girar la carretilla de manera que primero midiera del lado oriente y después el occidente. El resultado fue que la intensidad del occidente era 10% mayor que la del oriente. La conclusión indicó que la radiación cósmica primaria está constituida principalmente por partículas positivas, esto es, protones o núcleos atómicos, y no electrones.
En enero de 1933 fue publicado un artículo de Lemaître y Saldoval Vallarta con los resultados básicos de la teoría. Además del efecto de asimetría este-oeste, la teoría indicaba que si la carga eléctrica de las partículas de la radiación cósmica es predominantemente positiva, la intensidad medida en la dirección norte debería ser mayor que la medida en la dirección sur en el mismo ángulo cenital. El primero que sometió esta deducción teórica al experimento fue Thomas H. Johnson. Primero en Copilco, en el Distrito Federal, y después en el Nevado de Toluca, Johnson logró comprobar la existencia del efecto de asimetría norte-sur.
Estos experimentos cruciales tuvieron enorme resonancia, pues al determinar el signo de la carga eléctrica de las partículas se pudo identificarlas con precisión, es decir, se pudo comprender la naturaleza de la radiación cósmica.
Mientras Sandoval Vallarta y Lemaître trabajaban en su teoría, en varios países muchos otros científicos buscaban también la solución del problema.
En 1932, W. Heisenberg, en Alemania, a partir de un argumento puramente cualitativo, había hecho notar la posibilidad de que ocurriera una asimetría este-oeste en la intensidad de la radiación cósmica.
En el mismo año, Enrico Fermi y Bruno Rossi, del Instituto de Física de Florencia, en Italia, hicieron un estudio teórico en el que, como Sandoval Vallarta y Lemaître, encontraron que el uso del teorema de Liouville simplifica el análisis de los efectos geomagnéticos en la radiación cósmica.
Animado por este resultado y por los obtenidos en trabajos anteriores, Rossi trató de hallar una prueba experimental de la naturaleza de la radiación cósmica. En la hermosa colina de Arcetri, a corta distancia de la villa donde Galileo había pasado los últimos años de su vida como exiliado político, Rossi intentó detectar la asimetría este-oeste en un experimento sin éxito. Al analizar los resultados, Rossi se dio cuenta de que el efecto sería más pronunciado en un lugar con una latitud más baja y a una gran altura sobre el nivel del mar. De inmediato empezó a hacer planes para una expedición a Asmara, en Eritrea, que se encuentra a 11° 30’ de latitud norte y a dos mil trescientos sesenta metros de altura sobre el nivel del mar. Sergio de Benedetti, quien años más tarde se distinguiría como físico nuclear, se unió a la empresa. Los preparativos les tomaron un buen tiempo; cuando finalmente se encontraban dispuestos para emprender el viaje recibieron una desagradable sorpresa. El propio Rossi, después de cincuenta años, lo relata así: “Aquí, debo admitir que sufrimos una dolorosa decepción cuando encontramos que por unos meses habíamos perdido la prioridad de este importante descubrimiento. Ocurrió que, cuando ya estábamos listos para salir en nuestra expedición, leímos en The Physical Review dos artículos, uno escrito por Thomas Johnson, y otro, por Luis Álvarez y Arthur Compton, en los que se daba cuenta de la observación hecha en la ciudad de México y en una montaña cercana llamada Nevado de Toluca, del efecto de asimetría este-oeste”.
A pesar de no tener ya el aliciente de ser los primeros, pues Álvarez, Compton y Johnson se les habían adelantado, Rossi y De Benedetti prosiguieron con sus planes. En el otoño de 1933 llegaron al África Oriental y montaron su experimento en una cabaña ubicada en una colina cerca de Asmara. Pronto encontraron que la intensidad de los rayos cósmicos en la dirección occidental era considerablemente mayor que en la dirección oriental, confirmando así que la radiación cósmica primaria está constituida, por lo menos en parte, de partículas cargadas eléctricamente, y que la carga de estas partículas es positiva. Ese resultado, que ratificó el hallazgo de Álvarez, Compton y Johnson, sorprendió a Rossi y a De Benedetti, quienes, al igual que muchos otros físicos, esperaban que la radiación cósmica primaria estuviera formada principalmente de electrones, cuya carga eléctrica es negativa.
Las predicciones teóricas de Sandoval Vallarta y Lemaître, y su inmediata confirmación experimental llevada a cabo por Álvarez, Compton y Johnson, ponían a los físicos estadounidenses a la cabeza de una reñidísima competencia en la que participaban los mejores científicos de los países avanzados de Europa, América y Asia: C. Powell, J. G. Wilson y H. Elliot, de la Gran Bretaña; P. Auger y Le Prince-Ringuet, de Francia; E. Fermi, B. Rossi y S. Benedetti, de Italia; W. Bothe, W. Kolhörster, W. Heisenberg y su grupo, de Alemania; J. Clay y H. P. Berlage, de Holanda; C. Störmer, de Noruega; H. Yukawa y S. Kikuchi, de Japón, y muchos otros.
En el verano de 1932, el descubrimiento del efecto de latitud había sido presentado en México por Compton en una sesión combinada de las tres sociedades de ciencias mexicanas. La Sociedad Alzate, la Sociedad de Geografía y Estadística y la Sociedad de Ingenieros y Arquitectos, que así gozaron de las primicias de este hallazgo. Al año siguiente, en 1933, Sandoval Vallarta explicó en la Sociedad Alzate la teoría del efecto de latitud y la predicción de las asimetrías este-oeste y norte-sur, que se observarían en la intensidad de la radiación cósmica. En el verano de 1934 vino a México Thomas H. Johnson, quien en compañía de Manuel Sandoval Vallarta midió nuevamente la asimetría este-oeste en la ciudad de México, en Veracruz y en San Rafael (en la Caja de la Cuesta), confirmando los resultados de las medidas de Álvarez. En 1934, Johnson, Fussell y Sandoval Vallarta midieron la asimetría norte-sur en Copilco y en el Nevado de Toluca. Los resultados de estas últimas mediciones resolvían en favor de la teoría de Lemaître y Sandoval Vallarta la polémica planteada por el célebre físico teórico noruego Carl Störmer en una carta a Nature de 1933.
Durante las vacaciones de verano en la ciudad de México, Manuel Sandoval Vallarta, cuidadoso de no perder el contacto con los estudiosos de la física y las matemáticas de México, visitó a don Sotero Prieto, su antiguo maestro de matemáticas. En esos días Sandoval Vallarta presentó los resultados de sus trabajos en el Seminario de Física y Matemáticas que Prieto presidía en la Escuela de Altos Estudios. En ese seminario se presentaron y discutieron las predicciones y los resultados de la teoría de Sandoval Vallarta y Lemaître, así como la confirmación de los efectos de asimetría de la radiación cósmica y su verificación observacional. Sandoval Vallarta también asistió a las sesiones de la Sociedad Científica José Antonio Alzate, en donde presentó los resultados de sus investigaciones. Ahí ofreció, en conferencias formales, sus trabajos tanto de la teoría de la relatividad como de la mecánica ondulatoria. De esa manera, proporcionó a los investigadores y alumnos mexicanos una visión de los problemas de la física teórica de interés en ese momento con la profundidad y la actualidad propias del investigador de la máxima calidad académica que participa con éxito en su solución. Las pláticas de Sandoval Vallarta avivaron extraordinariamente el interés ya existente por el estudio de la física teórica, pero, sobre todo, su ejemplo vivo entusiasmó a algunos de sus miembros más jóvenes a convertirse en científicos profesionales como él. En ese grupo estaban, entre otros, Carlos Graeff Fernández, Nabor Carrillo y Alberto Barajas.
No es, pues, sorprendente, que en ese año de 1934 Sandoval Vallarta recibiera a Alfredo Baños, su primer estudiante mexicano, quien acudía al Instituto Tecnológico de Massachusetts para hacer un doctorado en Física bajo su dirección, y que, dos años más tarde, en 1935, se presentara el segundo, Carlos Graeff Fernández, que iba, al igual que Baños, a realizar un doctorado de Física bajo su dirección. Ambos se interesaban en contribuir al desarrollo de la teoría de los efectos geomagnéticos en la radiación cósmica.
Mientras tanto, en la Universidad Nacional Autónoma de México las jefaturas de grupo de Física y Matemáticas —que habían estado a cargo de don Basiliso Romo y don Sotero Prieto, respectivamente— habían sido fusionadas, en 1934, en la Facultad de Ciencias Físicas y Matemáticas, que reunía a las Escuelas Nacional de Ingenieros y Nacional de Química con el Departamento de Física y Matemáticas. Después de varias reformas universitaria, el 1 de marzo de 1937, se formó la Escuela Nacional de Ciencias Físicas y Matemáticas, que en 1938 se transformó en la Facultad de Ciencias. En 1938 se fundó el Instituto de Física y Matemáticas, cuya vida fue efímera, pues en noviembre de ese mismo año se disoció en un Instituto de Física y en otro de Matemáticas. El 1 de febrero de 1939 se aprobó oficialmente la formación del Instituto de Física de la UNAM, y se designó como su primer director a Alfredo Baños, quien había regresado a México ese año después de obtener el grado de doctor en Ciencias en el Instituto Tecnológico de Massachusetts, bajo la dirección de Manuel Sandoval Vallarta. En ese Instituto, Baños, con la colaboración de Héctor Uribe, Manuel L. Perusquía y Jaime Lifshitz, continuó las investigaciones sobre la dinámica del movimiento de las partículas cargadas en el campo magnético terrestre que había iniciado con Manuel Sandoval Vallarta. Ese grupo empezó de inmediato a llevar a efecto el programa de colaboración científica que Baños había formulado con el grupo del Instituto Tecnológico de Massachusetts que encabezaba Sandoval Vallarta. En el programa de colaboración se preveía la construcción y el montaje de un sistema detector de cuatro trenes de contadores Geiger-Müller para la medición del efecto azimutal y la distribución de la energía de la radiación cósmica primaria por el método propuesto por Sandoval Vallarta. Además, Baños, Lifshitz y Uribe estudiaron algunos problemas de la teoría del movimiento de partículas cargadas en el campo de un dipolo magnético y su aplicación a la teoría de los efectos geomagnéticos en los rayos cósmicos.
En febrero de 1942 fue inaugurado el Observatorio Astrofísico Nacional de Tonantzintla, Puebla y el mismo día se iniciaron ahí los trabajos del xvii Congreso Interamericano de Astrofísica, en donde Baños presentó los primeros resultados de las mediciones de la variación azimutal de la intensidad de la radiación cósmica a varios ángulos cenitales. Jaime Lifshitz y Héctor Uribe presentaron sus repectivos trabajos sobre la dinámica de las órbitas periódicas simétricas de las partículas cargadas en el campo mágnético terrestre. A ese congreso asistieron un buen número de científicos de primera línea, algunos de los cuales, como H. Shapley, S. Lefshetz y G. D. Birkhoff, tendrían una influencia duradera en el desarrollo de la física y las matemáticas en nuestro país.
De 1932 a 1939 los esfuerzos de Manuel Sandoval Vallarta, en colaboración con Georges Lemaître y los numerosos alumnos de ambos, habían estado encaminados a desarrollar la teoría de los efectos geomagnéticos en la radiación cósmica, confirmando así la teoría con numerosísimas observaciones. Esta es mundialmente conocida como la teoría Lemaître-Vallarta de la radiación cósmica.
En 1939, Manuel Sandoval Vallarta fue nombrado profesor titular de Física en el Instituto Tecnológico de Massachusetts, cargo que desempeñó hasta 1946.
En 1943 se inició lo que podría llamarse la época mexicana de Sandoval Vallarta como investigador científico. En ese año, el presidente Ávila Camacho formó la Comisión Impulsora y Coordinadora de la Investigación Científica con el propósito de atraer a México a don Manuel, al nombrarlo presidente y vocal físico-matemático de esta institución. Los viajes de Sandoval Vallarta a la ciudad de México se hicieron más frecuentes y sus estancias en esta capital más prolongadas. La participación de Estados Unidos en la Segunda Guerra Mundial trajo como consecuencia la reorientación de una fracción muy considerable de la organización científica de ese país hacia proyectos bélicos; en especial, el laboratorio del integrador diferencial de Vannevar Bush —la máquina de cómputo más avanzada de ese tiempo— fue dedicado a proyectos militares, lo que interrumpió el activísimo programa de cálculo de las órbitas de los rayos cósmicos que realizaba don Manuel con sus alumnos. Así fue como, en 1943, se inició en la ciudad de México, con la supervisión de Sandoval Vallarta, un programa de experimentos con un aparato diseñado, construido y calibrado por A. Baños y M. L. Perusquía. Este consistía de cuatro trenes de contadores Geiger-Müller dispuestos para registrar eventos a ángulos zenitales de 0, 20, 40 y 60 grados. Los contadores habían sido fabricados en el laboratorio de R. D. Evans en el Instituto Tecnológico de Massachusetts y estaban en la azotea del viejo Palacio de Minería, colocados en un cuarto muy pequeño llamado pomposamente la Torre del Contador, el cual operaba automáticamente y registraba fotográficamente el número de eventos cada treinta y dos minutos. Se tomaron lecturas en corridas ininterrumpidas que duraban por lo menos cien días, de julio de 1943 a mayo de 1946. El análisis estadístico de los datos fue hecho por Juan B. de Oyarzábal y la interpretación de éstos por Sandoval Vallarta, quien mediante un procedimiento semejante, años más tarde interpretó los resultados de las primeras medidas de la radiación cósmica hechas fuera de la atmósfera por Van Allen, con cohetes, en 1949. Al grupo de investigadores teóricos, en ese mismo año se agregó Ruth Gall.
No hay duda de que Manuel Sandoval Vallarta fue quien con su ejemplo atrajo hacia la investigación científica en la física teórica y experimental a quienes formaron los primeros grupos serios que trabajaron en México en este siglo. No hay duda, tampoco, de que fue él quien organizó, orientó y condujo a los éxitos iniciales a los primeros grupos teóricos y experimentales que lograron hacer buena investigación científica en física en México. Sin embargo, en la tarea de crear un nuevo paradigma del científico, reiniciar la tradición científica y echar andar de nuevo la física en México, hizo una labor aún más importante. Durante veintinueve años presidió el Seminario de física más importante del país. En 1948, en las instalaciones de la Comisión Impulsora y Coordinadora de la Investigación Científica, se volvió a reunir el Seminario de Física Teórica que había suspendido sus sesiones en 1935 por la muerte de Sotero Prieto, su principal animador. El único miembro del seminario de Prieto que participaba en el nuevo seminario fue don Manuel Sandoval Vallarta.
Todos los viernes, a las 17:15 en punto, después de presentar al orador en turno, se sentaba en el fondo del salón para poder seguir con atención la exposición y la discusión sin perder detalle. Al final hacía una resumen conciso y, con frecuencia, un comentario. Sus extensas relaciones personales permitieron que por el Seminario desfilaran los mejores físicos del mundo. Ponía el mismo interés y cuidado en seguir la exposición de un premio Nobel que la de un estudiante graduado que presentaba los resultados de su tesis. Casi todos los trabajos de física que se hicieron en México en esa época fueron presentados, discutidos y criticados en ese seminario, sobre todo en los primeros años. Fue ahí donde con sus comentarios, siempre comedido y cortés, Sandoval Vallarta fijó las normas de calidad académica que ahora rigen. En don Manuel, la amabilidad y la cortesía no estaban reñidas con el rigor científico y un agudo sentido crítico que le permitían hacer un juicio certero y exacto formulado en palabras breves y precisas, que, cuando era negativo, tenían un efecto demoledor. De igual modo era parco en los elogios, que aun siendo pocos y breves, eran siempre recibidos con agrado y satisfacción. Se puede afirmar, sin ninguna exageración, que la mayor parte de la física seria que se hizo o se hace en el país no hubiera podido realizarse si no se hubieran fijado oportunamente los estrictos criterios de calidad científica que estableció don Manuel a través de su seminario. El seminario, que se reunió primero en la Comisión Impulsora y Coordinadora de la Investigación Científica, luego en el Instituto Nacional de la Investigación Científica, después en el Instituto Nacional de la Investigación Científica, y más tarde en la Comisión de la Energía Nuclear y en el Instituto Nacional de Energía Nuclear, se reúne desde 1978 en el Instituto de Física de la unam, el mismo día y a la misma hora. El seminario lleva por nombre: Manuel Sandoval Vallarta.
|
||||
|
Alfonso Mondragón
Instituto de Física,
Universidad Nacional Autónoma de México.
|
||||
|
Simposio de rayos cósmicos en la Universidad de Chicago en 1939
|
||||
|
En esa época los mejores físicos del mundo tenían puesta la vista en la radiación cósmica. Antes de la década de 1940 no había aceleradores de partículas que pudieran dar información sobre la estructura interna de la materia. La única información sobre colisiones de alta energía, y por lo tanto de la composición de la materia, se podía obtener de las observaciones de los rayos cósmicos. Los datos que se obtenían a partir de los estudios de los rayos cósmicos únicamente pudieron ser interpretados a partir de la formulación de la teoría de Lemaître-Vallarta, que permitió el signo de la carga y el espectro de enrgías de las partículas primarias. En la radiación cósmica o asociada a ella se descubrieron el positrón (e+), el muión (m), los piones (p+,p-), las partículas cargadas y muchas otras.
Además del interés en la estructura de la materia, la radiación cósmica era interesante por sí misma, por la información astrofísica que podía dar, por los efectos geomagnéticos y su relación con la geofísica y por muchas otras razones. Así pues, interesaba por igual a los físicos experimentales, así como teóricos, a aquellos interesados en la estructura de la materia (Heisenberg, Fermi, Wheeler y otros), astrofísicos, geofísicos y otros curiosos.
En la primera fila, 2º de izquierda a derecha, Alfredo Baños, primer director del Instituto de Física y Jefe del Departamento de Física de la Facultad de Ciencias. En la tercera posición (12), Manuel Sandoval Vallarta.
Entre los participantes se encuentran algunas de las figuras más notables de la física, tanto teórica como experimental, de este siglo: 1. Hans Bethe, premio Nobel, de los fundadores de la Física Nuclear, formuló las bases de la Astrofísica Nuclear y formuló el ciclo (de y Bethe) de reacciones nucleares que dan calor al sol y la luz. 4. Arthur Holy Compton, premio Nobel por el efecto que lleva su nombre, la dispersión de fotones por partículas cargadas. 5. E. Teller, Premio Nobel, desarroló la física básica de la función nuclear y participó en el diseño de la bomba H. 8. S. Goudsmit, físico holandés que junto con Uhlenheck descubrió el spin del electrón, por lo que recibieron el premio Nobel. 11. J. R. Openheimer, encabezo el grupo de físicos que desarrolló la bomba atómica en Los Alamos. Además contribuyó de manera importante al desarrollo de la física de las partículas elementales y la teoría del campo cuántico. 12. Carl D. Andersen ganó el premio Nobel por el descubrimiento del positrón en una observación de la radiación cósmica. 15. Victor Hess, descubridor de la radiación cósmica por lo que recibió el premio Nobel. 17. Brunno Rossi, discípulo de Enrico Fermi, confirmó junto con Sergio de Benedetti el descubrimiento del efecto latitud que habían hecho Clay y Compton. Dedicó su vida a la investigación experimental y teórica de la radiación cósmica. 18. Wilhelm Bothe ganó el premio Nobel por sus contribuciones a la Física y a la Química Nuclear. Hizo el experimento crucial que sugirió que la radiación cósmica podría estar compuesta de partículas cargadas de electricidad. 19. Werner Heisenberg ganó el premio Nobel por el desarrollo de la mecánica cuántica en su formulación matricial. 20. Pierre Auger descubrió los chubascos de partículas en la radiación cósmica secundaria. 23. J. Clay de Amsterdam descubrió, simultáneamente a A. H. Compton, el efecto de latitud en la radiación cósmica. Hizo sus observaciones en Indonesia y el Pacífico sur. |
||||
|
_______________________________________________________________ como citar este artículo →
Mondragón, Alfonso. (1999). Manuel Sandoval Vallarta y la física en México. Ciencias 53, enero-marzo, 32-39. [En línea] |
||||

| ← Regresar al índice ⁄ artículo siguiente → |
 |
 |
|||
| de lo solube y lo insoluble |
||||
|
Reflexiones en torno al binomio
ciencia-sociedad
|
|
|||
|
Antonio R. Cabral
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
Ya es un lugar común decir que, en los últimos cincuenta años, la ciencia ha influido a la sociedad más que en toda su historia anterior. La biología molecular, en otro tiempo considerada como tema de novelas, es ahora uno de los motores más eficaces de la ciencia moderna. La ingeniería genética, la clonación de ovejas, los análisis de ácidos nucleicos depositados en vestidos azules son noticias de ocho columnas. Por todos lados se nota el impacto de la ciencia en la colectividad; esto es tan obvio que parece ya no haber nada más qué decir. La ciencia, y su sucedánea, la tecnología, son parte de prácticamente todas las actividades humanas, ya que de una u otra manera están inmersas no sólo en las casas que habitamos, en los alimentos que comemos y en los transportes que abordamos, sino también en los artículos electrónicos que usamos para trabajar o para no aburrirnos. Podría decirse que en mayor o menor grado, nadie escapa del arrope de la ciencia. Por ello, ahora que al parecer el planeta se ha librado de la destrucción nuclear, la sociedad presta atención a otros peligros atribuidos a la ciencia como la clonación de humanos, la creación ad libitum de virus tan temidos como letales y la manufactura de órganos a destajo. Los científicos podrían a su vez ser vistos como seres misteriosos que, metidos en sus laboratorios, maquinan ideas perversas de cómo dañar a sus semejantes sin dejar la menor huella; puede que esto no sea totalmente falso: no sería difícil que alguien con conocimientos profundos sobre la vida misma y que al mismo tiempo padezca debilidades humanas, quiera aplicar esos conocimientos para hacer el mal. Tampoco debe dudarse de la existencia de científicos que prefieran seguir otros caminos que los señalados por la ética científica, que no es otra que la ética cotidiana, y utilicen su experiencia para beneficio propio sin que medie el más mínimo interés en buscar la verdad por la verdad misma. En un mundo tan competitivo como el que ha tocado vivir no deben extrañar esos desvíos: los científicos, como los empresarios, los políticos, los choferes, los taxistas, los bibliotecarios, los sacerdotes y el público en general, nunca han pertenecido a la especie que tiene un nimbo sobre sus cabezas.
Con todo y las innovaciones tecnológicas, las asimetrías sociales están más presentes que nunca: el paludismo sigue matando a varios miles de personas al año en países africanos a pesar de que, en 1902, Sir Ronald Ross recibió el Premio Nobel de Medicina precisamente por sus descubrimientos sobre el ciclo vital del parásito causante de esa enfermedad. Igualmente, en las sociedades avanzadas una gran proporción de las riquezas nacionales se gastan en salud y recreación, pero en el mundo subdesarrollado las hambrunas y las guerras matan a millones de personas, la desnutrición y las enfermedades parasitarias son muy frecuentes y ni siquiera se pueden satisfacer las necesidades más básicas como la comida y la habitación. Desde luego que a la ciencia le gustaría decir que ya tiene la solución para la pobreza y que pronto todos los habitantes de la Tierra tendrán alimentos sobre sus mesas tres veces al día. Todos sabemos que estos problemas, por ser del dominio de los gobiernos y de su política económica, rebasan los límites de la ciencia, y aun los de la economía, la cual, dicen sus practicantes, es parte de ella. Pero no por esto, la sociedad debe sentirse defraudada; al contrario, debe estar segura de que la ciencia, al hacerse preguntas que pertenecen al mundo real, tarde o temprano, encontrará respuestas que tengan una aplicación práctica. En la mayoría de los países la investigación científica está patrocinada principalmente por agencias gubernamentales, ya que la participación de la industria privada en los países subdesarrollados, con algunas excepciones, es magra. La sociedad analiza los logros de los hombres de ciencia mediante comisiones evaluadoras de la actividad científica y confía en que el dinero destinado para ellos es bien utilizado. En los países desarrollados, el gasto asignado a la investigación científica se escatima cada vez menos, pues los gobernantes saben que esto trae consigo bienestar y riqueza para los ciudadanos y con ello, hay que decirlo, la posibilidad de reelegirse; pero en los países menos favorecidos, la sociedad aún no se ha dado cuenta de que la ciencia es un vehículo seguro para salir del subdesarrollo y no al revés. Según lo apuntó nuestro admirado Voltaire, esto no sorprende: es difícil que la razón prevalezca ahí donde domina la superstición. Mientras en Estados Unidos el número de científicos por cada diez mil habitantes es de treinta y cinco, en México es de uno. No debemos imitar el camino andado por los estadunidenses porque sea el mejor modelo, sino porque, con todo y sus limitaciones, la ciencia ofrece alternativas razonables, y para, como dice el doctor Ignacio Chávez, “no pasarnos la vida rezando las verdades y los errores que nos legaron otros”.
El quid no es la falta de creatividad, ya que los científicos que habitan el Tercer Mundo no son cortos de ideas, sino que padecen la enfermedad que el biólogo molecular inglés avecindado en California, Sydney Brenner, reciente y atinadamente bautizó como mdd, que en nuestro idioma bien podría traducirse con puntualidad como “Millones de Dólares”, pero que en el de él es “Money Deficiency Disease”. A nadie conviene padecer la enfermedad de Brenner; cuando ésta ataca sólo a una persona, las secuelas no son muy graves porque no afectan directamente a la sociedad. En cambio, cuando la ciencia y la tecnología de un país enferman de mdd, los síntomas rebasan al paciente y sus repercusiones son incalculables. Dinero y bienestar están íntimamente ligados, no en vano el doctor Amartya Sen recibió este año el Premio Nobel de Economía por sus estudios sobre la calidad de vida, concepto éste que pocos entienden y que nadie sabe definir, pero que pocos pueden rebatir. El impacto de la ciencia en la colectividad, ya quedó dicho, es indiscutible. Desde hace algunos años diversos científicos estadunidenses intentan convencer a sus compatriotas para que dejen entrar la ciencia en sus modelos de pensamiento y, consecuentemente, en su acción. Ya se verán los efectos. En nuestro país hay esfuerzos en el mismo sentido, aunque, seguramente, son insuficientes. La afrenta es doble: producir a pesar de la mdd e incrementar la cultura científica de un país en donde la ciencia no es un valor.
|
||||
|
Antonio R. Cabral
Instituto Nacional de la Nutrición “Salvador Zubirán”.
_______________________________________________________________ como citar este artículo →
Cabral R., Antonio.(1999). Reflexiones en torno al binomio ciencia-sociedad. Ciencias 53, enero-marzo, 14-15. [En línea]
|
||||
|
|
||||