
")

Energía
 |
||
|
Ana María Sánchez, María Trigueros y
Julia Tagüeña
Historias de la ciencia y la técnica,
UNAM, México, 1999.
|
||
| HTML ↓ | ← Regresar al índice | |
|
Energía... Cuántas veces hemos oído esta palabra. Aunque la usamos muy a menudo, ¿podríamos decir qué es?
La energía es un concepto complejo de definir pero muy útil. Lo utilizan casi todas las ciencias y es también parte de nuestro vocabulario cotidiano.
¿Hace cuánto tiempo que hablamos de energía? Estamos tan habituados a hacerlo que pensamos que el concepto es muy antiguo, pero no es así. Los antiguos griegos usaban la palabra “energía” para referirse a la eficacia, el poder o la virtud para obrar. Ni Galileo ni Newton, siglos después, conocían el concepto en la forma en que hoy lo manejamos. No fue sino hasta mediados del siglo xix cuando varios científicos que hacían experimentos diferentes en diversos lugares encontraron que fenómenos que hasta entonces se pensaban ajenos unos a otros, como el calor y el movimiento, la electricidad y el magnetismo, el movimiento y la luz, la afinidad química y el calor, y otros más, se relacionaban entre sí. Es más, estos fenómenos podían transformarse uno en el otro y en esa transformación podía definirse un concepto abstracto que da cuenta de algo que se conserva: la energía.
La energía es un concepto unificador en la naturaleza, y por su carácter abstracto es más fácil de definir en términos de sus manifestaciones.
La energía se manifiesta de muchas formas que pueden intercambiarse: mecánica, calorífica, eléctrica, química, magnética, nuclear. Las distintas formas de energía se transforman unas en otras, y lo más importante es que en ese proceso la cantidad total de energía se conserva. A este fenómeno se le llama principio de conservación de la energía.
El descubrimiento del principio de conservación de la energía representó un momento importante en la historia de las ciencias. Su nacimiento permitió descubrir que varias de ellas, que hasta ese momento se estudiaban por separado, constituían partes de un todo: la física. Desde su nacimiento este principio ha estado ligado íntimamente a la tecnología. Su aplicación permitió el desarrollo de nuevas industrias que cambiaron totalmente nuestra forma de vida.
El concepto de energía ha sido un motor del desarrollo de la ciencia y la tecnología. Su papel ha sido fundamental y sólo mediante un conocimiento profundo de este concepto y de sus consecuencias podremos comprender y manejar los efectos de la tecnología sobre el ambiente y la sociedad.
_______________________________________________________________
como citar este artículo → Sánchez, Ana María y Trigueros María, Tagüeña Julia. (1999). Energía. Ciencias 53, enero-marzo, 70. [En línea]
|
||
|
|
← Regresar al índice | |
El método de las ciencias
 |
||
|
Rosaura Ruiz y Francisco J. Ayala
Fondo de Cultura Económica, México, 1998.
|
||
| HTML ↓ | ← Regresar al índice / artículo siguiente → | |
|
La ciencia se mueve, cambia de lugar, se agita. Es veleidosa y frágil. Por dentro y por fuera, se transforma. Está tan viva como un ejemplar de una especie, libre o en cautiverio, que muestra facetas desconocidas aun para los mismos que, en apariencia, la elaboran con absoluto control, y se les escapa, huye. Y luego se queda quieta, inmóvil, camuflada, a la espera, como si supiera que una infinidad de miradas la acechan, pendientes de ella, para aprehenderla. Los científicos saben todo esto y saben también que la certeza absoluta no es inherente al método científico, no es infalible; avanza cuando unos científicos refutan teorías de otros científicos, o de sí mismos al reconocer equivocaciones, y entonces aquél adquiere uno de sus rasgos más significativos, el lado humano, en el momento en que hace que la ciencia avance, socializada por vía del lenguaje. He aquí el tema central de esta obra de Ruiz y Ayala: a la ciencia se la puede estudiar y comprender de igual modo que como han procedido las epistemologías evolucionistas, sólo que en este caso no son los entes vivos la unidad de conocimiento, sino los conceptos y las teorías, en su origen, devenir y futuro, partiendo de la idea de que todo términos darwinistas, aunque por supuesto, en la aproximación de este problema es necesario adecuar el darwinismo a la teorización de carácter social, ya que los primeros darwinistas también pensaron que su paradigma era tan general que podía extenderse a toda entidad cambiante. Así, las maneras discrepantes en que varias teorías tratan de dar cuenta de determinado problema —específicamente el desarrollo de la ciencia, con sus controversias y polémicas— se analizan en estas páginas en las que el propósito más importante de los autores es estimular el interés en estas teorías, reconocer sus fundamentos, identificar lo que hay de válido en ellas y definir sus límites.
_______________________________________________________________
como citar este artículo → Ruiz Gutiérrez, Rosaura y Ayala, Francisco J. (1999). El método de las ciencias. Ciencias 53, enero-marzo, 69. [En línea]
|
||
|
|
← Regresar al índice / artículo siguiente → | |
|
El diablo de los números
|
 |
|
|
Hans Magnus Enzensberger
Ediciones Siruela, España, 1998
|
||
| HTML ↓ | ← Regresar al índice / artículo siguiente → | |
|
Hacía mucho que Robert estaba harto de soñar. Se decía: Siempre me toca hacer el papel de tonto.
Por ejemplo, en sueños le ocurría a menudo ser tragado por un pez gigantesco y desagradable, y cuando esta a punto de ocurrir llegaba a su nariz un olor terrible. O se deslizaba cada vez más hondo por un interminable tobogán. Ya podía gritar cuanto quisiera ¡Alto! o ¡Socorro!, bajaba más y más rápido, hasta despertar bañado en sudor.
A Robert le jugaban otra mala pasada cuando ansiaba mucho algo, por ejemplo una bici de carreras con por lo menos veintiocho marchas. Entonces soñaba que la bici, pintada en color lila metálico, estaba esperándole en el sótano. Era un sueño de increíble exactitud. Ahí estaba la bici, a la izquierda del botellero, y él sabía incluso la combinación del candado: 12345. ¡Recordarla era un juego de niños! En mitad de la noche Robert se despertaba, cogía medio dormido la llave de su estante, bajaba, en pijama y tambaleándose, los cuatro escalones y... ¿qué encontraba a la izquierda del botellero? Un ratón muerto. ¡Era una estafa! Un truco de lo más miserable.
Con el tiempo, Robert descubrió cómo defenderse de tales maldades. En cuanto le venía un mal sueño pensaba a toda prisa, sin despertar. Ahí está otra vez este viejo y nauseabundo pescado. Sé muy bien qué va a pasar ahora. Quiere engullirme. Pero está clarísimo que se trata de un pez soñado que, naturalmente, sólo puede tragarme en sueños, nada más. O pensaba: Ya vuelvo a escurrirme por el tobogán, no hay nada que hacer, no puedo parar de ningún modo, pero no estoy bajando de verdad.
Y en cuanto aparecía de nuevo la maravillosa bici de carreras, o un juego para ordenador que quería tener a toda costa —ahí estaba, bien visible, a su alcance, al lado del teléfono—, Robert sabía que otra vez era puro engaño. No volvió a prestar atención a la bici. Simplemente la dejaba allí. Pero, por mucha astucia que le echara, todo aquello seguía siendo bastante molesto, y por eso no había quien le hablara de sus sueños.
Hasta que un día apareció el diablo de los números.
Robert se alegró de no soñar esta vez con un pez hambriento, y de no deslizarse por un interminable tobogán desde una torre muy alta y muy vacilante. En su lugar, soñó con un pradera. Lo curioso es que la hierba era altísima, tan alta que a Robert le llegaba al hombro y a veces hasta la cabeza. Miró a su alrededor y vio, justo delante de él, a un señor bastante viejo, bastante bajito, más o menos como un saltamontes, que se mecía sobre la hoja de acedera y le miraba con ojos brillantes.
—¿Quién eres tú?— preguntó Robert.
El hombre le gritó, sorprendentemente alto:
—¡Soy el diablo de los números!
Pero Robert no estaba de humor para aguantarle nada a semejante enano.
—En primer lugar —dijo—, no hay ningún diablo de los números.
—¿Ah, no? ¿Entonces por qué estás hablando conmigo, si ni siquiera existo?
—Y en segundo lugar, odio todo lo que tiene que ver con las Matemáticas.
—¿Por qué?
—“Si dos panaderos hacen 444 trenzas en seis horas, ¿cuánto tiempo necesitarán cinco panaderos para hacer 88 trenzas?” Qué idiotez —siguió despotricando Robert—. Una forma idiota de matar el tiempo. Así que ¡esfúmate! ¡Largo!
El diablo de los números se bajó con un elegante salto de su hoja de acedera y se sentó al lado de Robert, que en protesta se había sentado entre la hierba, alta como un árbol.
—¿De dónde te has sacado esa historia de las trenzas? Seguro que del colegio.
—¡Y de dónde si no! —dijo Robert—. El señor Bockel, ese principiante que nos da Matemáticas, siempre tiene hambre, a pesar de estar tan gordo. Cuando cree que no le vemos porque estamos haciendo los deberes, saca una trenza de su maletín y se la devora mientras nosotros hacemos cuentas.
—¡Vaya! —exclamó el diablo de los números, sonriendo con sorna—. No quiero decir nada en contra de tu profesor, pero la verdad es que eso no tiene nada que ver con las Matemáticas. ¿Sabes una cosa? La mayoría de los verdaderos matemáticos no sabe hacer cuentas. Además, les da pena perder el tiempo haciéndolas, para eso están las calculadoras. ¿No tienes una?
—Sí, pero en el colegio no nos dejan usarla.
—Ajá —dijo el diablo de los números—. No importa. No hay nada que objetar a un poco de práctica con las tablas. Puede ser muy útil si uno se queda sin pilas. ¡Pero las Matemáticas, ratoncito, eso es muy diferente!
_______________________________________________________________
como citar este artículo → Magnus Enzensberger, Hans. (1999). El diablo de los números. Ciencias 53, enero-marzo, 68. [En línea] |
||
|
|
← Regresar al índice / artículo siguiente → | |
 |
 |
|||||||||
| del tintero |
||||||||||
| Zoologías |
|
|||||||||
|
conoce más del autor
|
||||||||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||||||||
|
||||||||||
|
||||||||||
|
||||||||||
|
||||||||||
|
_______________________________________________________________ como citar este artículo →
Neruda, Pablo. (1999). Zoologías. Ciencias 53, enero-marzo, 64-67. [En línea] |
||||||||||




 |
 |
|||||
|
El lirio acuático, ¿una planta nativa
de México?
|
|
|||||
|
María Guadalupe Miranda A. y Antonio Lot Helgueras
conoce más del autor
|
||||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||||
|
El lirio acuático (Eichhornia crassipes) es una planta libre flotadora miembro de la familia de las Pontederiaceae, que ocupa un lugar sobresaliente entre las comunidades de hidrófitas de agua dulce de las regiones tropicales y subtropicales del mundo. Las flores que la conforman, por su forma y color, son tanto la estructura más vistosa de la planta como el principal motivo que ha favorecido su introducción en estanques y acuarios de todo el mundo. Los lóbulos de la flor presentan tonalidades que van del color morado al lila y al azul claro, destacando en el lóbulo superior central una mancha de color amarillo.
Su distribución en México es sumamente amplia, ya que crece en una gran diversidad de hábitats dulceacuícolas localizados desde el nivel del mar hasta los 2 250 metros de altitud. Los únicos lugares en los que no se ha registrado son los estados de Baja California Sur, Chihuahua, Zacatecas, Tlaxcala y Yucatán.
La abundancia y la frecuencia con la que encontramos el lirio acuático (Eichhornia crassipes) en los cuerpos de agua naturales y artificiales en México nos hace pensar que se trata de una planta nativa del país.
En las regiones fuera de su área natural de distribución, el lirio acuático es considerado maleza (planta indeseable) por la serie de problemas que acarrea su presencia y, sobre todo, por el crecimiento tan rápido de su población. En hábitats perturbados sustituye con frecuencia a elementos de la flora nativa. Tiene diversos efectos perjudiciales, entre los que se pueden mencionar el entorpecimiento de la navegación por ríos y lagunas, la elevación de los índices de evaporación y el impedimento del paso de la luz al fondo de los estanques, lo que trae como consecuencia la eliminación de microalgas que son el alimento de crustáceos y peces. Sabemos que el lirio acuático es una de las malezas, o planta invasora, más difíciles de exterminar en áreas inundables manejadas por el hombre en las regiones tropicales y subtropicales del mundo.
Según Orozco-Segovia y Vázquez-Yanes esta planta es considerada nativa de México y de Centroamérica, aunque existen dudas sobre las causas de su distribución fuera de las cuencas del Amazonas y del Orinoco, que corresponden al centro de origen y de distribución primaria, aunque estos autores también señalan que su presencia en México podría deberse a una introducción temprana.
En la búsqueda que realizamos en diversas fuentes botánicas antiguas del país hallamos algunos datos que parecen apuntar hacia la segunda suposición. En la Historia de las cosas de la Nueva España, obra escrita por Fray Bernardino de Sahagún, publicada cerca de 1590, se hace referencia a las plantas que existían en los lagos, pero no se incluye al lirio acuático. Igualmente, en la Historia natural de la Nueva España, publicada en 1576 por el naturalista Francisco Hernández, protomédico del rey Felipe II, se mencionan algunas plantas acuáticas, pero, una vez más, no al lirio acuático. En el Códice Badiano, escrito en 1552 reconocimos íconos de macrofitas acuáticas, pero el lirio acuático tampoco aparece.
Buscando las plantas útiles del México antiguo, Torres revisó en 1985 otras obras del siglo XVI, como las crónicas de José Acosta, Antonio de Ciudad Real, Fray Diego de Landa y Fray Alonso de Molina, en donde no se hace ninguna referencia al lirio acuático.
Algunos estudios palinológicos recientes que se han realizado en la zona central de México con objeto de encontrar registros de polen de lirio acuático en estratos correspondientes a épocas precolombinas no han tenido éxito. Todo esto nos hace pensar que el lirio acuático no formaba parte de la flora acuática de México en esa época.
En 1882, en el Calendario botánico del valle de México, Bárcena tampoco menciona al lirio acuático. El primer registro lo encontramos hasta 1903, en la obra de Manuel Urbina, después de la cual otros autores como Reiche, Ramírez-Cantú y Conzatti ya lo consideran parte de la flora acuática mexicana.
El origen
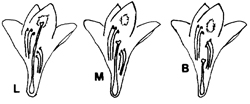
En el lirio acuático la morfología del órgano femenino de la flor o estilo varía en longitud y recibe los nombres de longistila (L), mesostila (M) y brevistila (B), respectivamente de mayor a menor talla (figura 1). A este fenómeno se le conoce como “heterostilia trimorfa” o “tristilia”.
Asumiendo que la mayor diversidad de formas de las especies se encuentra en la cercanía al centro de origen y de dispersión, y sin descartar la influencia del ambiente en la manifestación de la característica, podemos concluir que el lirio acuático es originario de la cuenca del Amazonas, en Brasil, ya que es únicamente allí donde se ha reportado la “tristilia”. También hay registros de “tristilia” en lagos cercanos a la confluencia de los ríos Paraguay y Paraná, en Argentina, en donde posiblemente se dispersó por medios naturales.
Las poblaciones de lirio acuático en Guyana, Venezuela y Colombia, así como en el Caribe, América Central y zonas templadas de América del Norte, parecen ser resultado de introducciones, ya que carecen del tamaño intermedio (la forma B). Esta idea se refuerza por la carencia de polinizadores especializados en América Central. El polinizador especializado registrado para el lirio es una abeja con lengua larga (Ancyloscelis gigas), que se ubica únicamente en la región amazónica donde se presenta la “tristilia”.
Resulta pertinente preguntarse si el lirio acuático pudo haber llegado a Centroamérica y a Norteamérica a través de mecanismos de distribución natural. Los órganos vegetativos de dispersión del lirio acuático incluyen las plantas completas u otras estructuras, como rizomas y estolones, y sólo pueden colonizar nuevos sitios que estén inundados. La dispersión es resultado del movimiento de los órganos vegetativos en los ríos, o bien, de las inundaciones que pueden acarrear los propágulos a lagos adyacentes, canales o diques.
Otro mecanismo de transporte de las semillas de algunas plantas acuáticas es el viento, pero las distancias alcanzadas son muy cortas. En el caso del lirio acuático, es poco probable que el viento las disperse, pues no tienen estructuras de transporte aéreo, como sucede con el tule ancho (Typha) y el carrizo (Phragmites).
Desde hace un siglo, Darwin explicó que la amplia distribución de ciertas especies vegetales de agua dulce se debía al acarreo de semillas y fragmentos vegetativos sobre el plumaje y las patas de las aves acuáticas. No obstante, para el caso del lirio acuático, que como ya mencionamos es una planta originaria de Brasil, es riesgoso considerar que este mecanismo como sea el responsable de las poblaciones del lirio asentados en Centro y Norteamérica, pues se carece de pruebas que indiquen su presencia desde el pasado.
Sin embargo, el hombre, de manera intencional, ha intervenido en la introducción y en la naturalización de nuevas plantas entre los hemisferios y entre los continentes. Sobre el lirio acuático, gracias a la magnífica obra compiladora de Gopal, se han documentado numerosos casos de introducciones a través de jardines botánicos instalados a partir de diversas exploraciones de naturalistas europeos en la región amazónica, en donde lo atractivo de su inflorescencia llamó considerablemente la atención.
Uno de los casos de introducción y naturalización del lirio acuático mejor conocidos sucedió en Norteamérica en 1884, con motivo de una exhibición de horticultura en la exposición de algodón en Nueva Orleans. Posteriormente, hacia 1894, en el delta del río Mississipi en Lousiana se registró el primer obstáculo serio para navegar en los ríos y corrientes que desembocaban en el golfo de México. Se considera que la naturalización del lirio acuático en esa región de Estados Unidos ocurrió durante la década de 1860 a 1870, y que hacia 1880 y, claramente después de 1900, el lirio acuático ya había invadido otras regiones como los Everglades en Florida. Más adelante, en 1903, el lirio se colectó por primera vez en Texas y, al año siguiente, ya se encontraba en California.
En Las chinampas del Distrito Federal, de Santamaría, se hace referencia a la introducción del lirio acuático a México: “entre las plantas acuáticas destaca el lirio acuático que es conocido desde hace solo catorce años, en que fue introducido por orden del general Carlos Pacheco, entonces secretario de Fomento”. De acuerdo con esta información, fue en 1898 cuando el lirio acuático llegó a los lagos del valle de México.
También debe considerarse la dispersión accidental, en particular de las semillas y de las fracciones vegetativas que crecen entre los cultivos irrigados y que se distribuyen con los productos cosechados y a través de embarcaciones, vehículos y maquinaria agrícola.
Su fácil dispersión
La dispersión del lirio y su establecimiento exitoso están directamente relacionados con las características ambientales abióticas y bióticas. A pesar de la amplia tolerancia a los factores abióticos del medio, y de ser una especie que puede llegar a invadir cuerpos de agua en regiones templadas y a altitudes considerables (su registro a mayor altitud en el mundo es en la ciudad de México), el lirio acuático no soporta las heladas y su crecimiento es limitado por la salinidad. En todo el mundo, su crecimiento óptimo se da a pH de 7 y adquiere el rango de infestación masiva entre 6.2 y 7.
En la variedad de hábitats de los sistemas fluviales del Amazonas y de los ecosistemas acuáticos de la región neotropical, en general, el lirio compite con otras especies de hidrófitas libres flotadoras (Azolla spp., Hydromistria stolo nífera, Pistia stratiotes, Lemna spp., Salvinia spp.) en mosaicos densos típicos de este tipo de comunidades naturales. Es interesante notar que el crecimiento masivo y con características de infestación de cualquiera de las especies mencionadas (especialmente Salvinia) en sus áreas naturales de distribución, sólo se ha registrado en donde artificialmente se cambió el régimen hidrológico, favoreciendo la formación de poblaciones dominadas por una sola especie.
La química del agua no limita el crecimiento del lirio acuático, pues su rango de tolerancia es amplio y el único factor que lo afecta es la carencia de fósforo. El lirio puede establecerse en aguas con pocos nutrientes, debido a las asociaciones microbianas que presenta en la rizosfera, y mientras más eutrófico sea el medio, más éxito tiene su propagación.
Entre las características de adaptación más importantes del lirio acuático están: a) su rápida capacidad de reproducción vegetativa; b) la habilidad para regenerarse a partir de pequeñas porciones del talo vegetativo; c) la completa o parcial independencia de la reproducción sexual; d) la morfología del crecimiento, que resulta en el desarrollo de la mayor área posible de tejido fotosintético, con relación al total de la planta y el cual rápidamente ocupa la superficie del agua, y e) independencia de las condiciones del sustrato y del nivel del agua.
Existen especies taxonómicamente cercanas, las cuales no son invasoras. Así, Eichhornia azurea es una especie estrechamente cercana a E. crassipes, con patrones de distribución nativa similar. Además, en contraste al hábito libre flotador de E. crassipes, E. azurea y las otras seis especies del género están unidas al sustrato. Es claro que la habilidad para llevar a cabo una propagación vegetativa rápida, unida al alto grado de movilidad, ha favorecido la dispersión de E. crassipes por encima de E. azurea.
Las fluctuaciones del nivel del agua inducen al lirio acuático a adoptar una forma emergente, con la elongación del pecíolo. Cuando la desecación es larga, los estolones quedan entrelazados esperando la época de inundación. A esta capacidad para modificar la forma y el tamaño de sus estructuras vegetativas cuando crecen bajo diferentes condiciones ambientales se le denomina plasticidad morfológica. Este atributo se ejemplifica bien en el lirio con la “heterofilia” o con la presencia en un solo individuo de dos o más tipos diferentes de hojas. La plasticidad morfológica y la habilidad para experimentar un crecimiento vegetativo rápido son los atributos clave que le dan a Eichhornia crassipes el éxito en la competencia interespecífica con otras plantas acuáticas invasoras.
Como en otras muchas macrofitas, en el lirio acuático hay una tendencia a reemplazar la reproducción sexual por la asexual. El ciclo completo de semilla a semilla es muy lento, ya que dura más de cinco meses. Además, para la germinación de la semilla es un requisito indispensable, la escarificación (rompimiento de la cubierta externa). En la propagación vegetativa el lirio produce estolones (tallos horizontales) que desarrollarán hojas arrosetadas de una yema terminal. Este proceso se repite en las plantas hijas y cuando se tienen las condiciones ideales, un número inmenso de plantas se pueden producir en corto tiempo, hasta duplicar la población en un periodo que va de cinco a quince días.
La presencia masiva e invasora del lirio ha llevado a buscar soluciones para su control. El hecho de que no existan herbívoros naturales, ni plagas que lo afecten, ha llevado a que actualmente se realicen estudios para utilizar hongos patógenos que puedan limitarlo. También se estudian diferentes artrópodos (coleópteros y lepidópteros) que puedan eliminarlo y se han probado, además, peces herbívoros como control biológico, entre los que podemos mencionar: Ctenopharyngodon idella, Tilapia rendalli y Metynnis roosevelti.
Una planta introducida
A través de los registros históricos, antropológicos y palinológicos, no se ha podido comprobar la existencia del lirio acuático en México en la época precolombina, por lo que se podría pensar que fue introducido voluntariamente como planta de ornato en Estados Unidos hacia 1884. Es posible que de ahí se haya dispersado en forma natural por medio de aves migratorias hasta llegar a nuestro país.
De cualquier forma, su introducción o naturalización en los lagos mexicanos no debió ocurrir antes de 1884. Lo que si es claro es su establecimiento y creciente poblamiento en nuevos espacios, desplazando a otras plantas acuáticas más sensibles a la perturbación humana.
La presencia del lirio acuático en ecosistemas lacustres estables con una alta biodiversidad es un fenómeno raro y su condición de planta “oportunista” sólo ocurre con la alteración del hábitat, especialmente cuando se presenta un incremento de nutrientes (eutroficación) por el derrame de fertilizantes o aguas negras. Desafortunadamente, hoy día es alarmante la modificación de los ambientes acuáticos naturales y el aumento de cuerpos de agua artificiales (presas, canales de irrigación, estanques de crecimiento, etcétera) que, por su inevitable conexión, pueden eventualmente contaminar grandes sistemas acuáticos a través de los ríos y de las crecientes durante los meses de lluvias y con la presencia de ciclones y otras perturbaciones meteorológicas. Con ello se favorece la invasión de Eichhornia crassipes como maleza a niveles que no pueden de controlar ni la naturaleza ni el hombre.
|
||||||
|
Referencias Bibliográficas
Gopal, B. 1987. Water Hyacinth. Aquatic Plant Studies. 1. Amsterdam, Elsevier Science Publishers, 471 p.
Mitchell, D. 1974. Aquatic vegetation and its use and control. Nueva York, unesco. pp. 38-49.
Orozco-Segovia, A. y C. Vázquez-Yáñez. 1993. Especies Invasoras: su impacto sobre las comunidades bióticas. México, Pronatura, A.C. pp. 29-31.
Sculthorpe, C. 1967. The biology of aquatic vascular plants. Londres, Edward Arnold. 610 p.
West, R. y P. Armillas. 1959. “Las chinampas de México, poesía, y realidad de los jardines flotantes”, en Cuadernos Americanos, 50, pp. 165-182.
|
||||||
|
María Guadalupe Miranda A.
UAM, Iztapalapa.
Antonio Lot Helgueras
Instituto de Biología,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Miranda A., María Guadalupe y Lot Helgueras, Antonio. (1999). El lirio acuático, ¿una planta nativa de México? Ciencias 53, enero-marzo, 50-54. [En línea] |
||||||
|
|
||||||
 |
 |
|||
| del bestiario |
||||
| La biodiversidad en el Asteroide B 612 |
|
|||
|
Héctor T. Arita
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
Si existiera un programa de conservación de la biodiversidad a nivel del sistema solar, el asteroide B 612 difícilmente se contaría entre las áreas prioritarias para establecer una reserva biológica. Efectivamente, si un hipotético exobiólogo realizara el inventario del planetoide hogar del Principito de Saint-Exupery, se encontraría con una flora representada por una vanidosa flor encerrada en un domo protector, unas cuantas flores de otra especie no muy bien determinada y unos cuantos brotes de baobab. Asimismo, encontraría que no existe fauna nativa en el lugar, y que el único animal es un borreguito encerrado en una caja. El asteroide no entraría, definitivamente, en las listas de sitios de megadiversidad.
El término “megadiversidad” fue acuñado hace algunos años por el biólogo estadounidense Russell Mittermeier para definir a la docena de países en los que se concentra la mayor parte de la diversidad biológica del planeta. Para identificar estos países megadiversos, Mittermeier recopiló información sobre el número de especies de los principales grupos biológicos y constató que existen unas pocas naciones que sobresalen en casi todos los rubros. México, por ejemplo, está incluido en este selecto grupo de países porque tanto su flora de plantas vasculares como sus faunas de reptiles, anfibios y mamíferos se encuentran entre las diez más ricas del planeta en número de especies. La riqueza de especies, es decir, el total de especies de plantas o animales que se encuentran en un sitio dado, es un criterio relativamente fácil de medir e interpretar, y en muchos casos se ha usado como sinónimo de biodiversidad. Para muchos biólogos, el conteo de especies se ha convertido en una especie de obsesión por cuantificar de alguna manera un fenómeno tan complejo como lo es la diversidad biológica. Un libro reciente sobre el tema, por ejemplo, define a la biodiversidad como “una biología de números y diferencias”.
El Principito no se preocuparía en lo más mínimo si un exobiólogo exageradamente cuantitativo menospreciara su pequeño asteroide debido a su pobreza de especies. Seguramente tal biólogo le haría recordar al hombre de negocios que vivía en otro asteroide y que había logrado contar quinientos millones seiscientos veintidós mil setecientas treinta y una estrellas y que se pensaba dueño de todas ellas por el simple hecho de haberlas catalogado. Podemos imaginar a un exobiólogo visitando el pequeño planetoide del Principito, observando con desdén a su alrededor y comentando en forma despectiva que en la Tierra existe un número aún no determinado de especies de plantas y de animales, pero que la cifra seguramente rebasa las treinta millones de especies. El Principito, sin inmutarse demasiado, se encogería de hombros y preguntaría: “¿Y qué hacen con todas esas especies?”, para rematar con una de sus frases favoritas: “Los adultos son definitivamente extraordinarios”.
Sin duda, la catalogación de las especies de plantas y animales de cada país es un paso importante para el conocimiento y el adecuado uso de la riqueza biológica. Sin embargo, esa labor debe ir aparejada por una comprensión más profunda de lo que implica el término biodiversidad. Hay que recordar que lo que ahora observamos como diversidad biológica es el resultado de millones de años de evolución orgánica y que las diferentes especies que observamos hoy día no son sino un pálido reflejo de la increíble variedad y variabilidad dentro y entre los organismos vivos. Cierto, el conteo de especies nos sirve de guía para estimar la diversidad de un sitio, pero poco nos dice sobre la variación que existe entre los individuos de cada una de las especies, además no toma en cuenta la diversidad genética contenida en esos organismos, ignora la estructura de las comunidades de plantas y animales y olvida la complejidad de las relaciones funcionales dentro de los ecosistemas.
El caso de México es muy ilustrativo. Sin duda es interesante e importante saber que en nuestro país existen alrededor de veinteséis mil especies de plantas vasculares, cerca de doscientos ochenta de anfibios, más de setecientas de reptiles y aproximadamente cuatrocientos cincuenta de mamíferos terrestres. Tal vez también es relevante conocer que nuestro país cuenta con treinta y cinco especies de esquizómidos, diez ricinuleidos, trecerafidiópteros y nueve mecópteros, pero si nos quedamos únicamente con los números fríos y no vemos más allá del simple conteo, caeríamos en el mismo absurdo que el hombre de negocios que se creía dueño de millones de estrellas y lo único que poseía era un papel en el que anotaba sus compilaciones estelares.
Dentro de cada especie, cada individuo es un reservorio único de información genética que se manifiesta en la variabilidad que presentan todas las especies de plantas y animales. Esta variabilidad se hace evidente a nivel local en las diferencias que existen entre los individuos de una población, y a nivel del país en la variación geográfica de las especies. Por ejemplo, todos los venados de cola blanca del país pertenecen a la misma especie (Odocoileus virginianus), pero no se necesita ser especialista en biodiversidad para percibir diferencias entre los venados de un sitio en particular: habrá algunos que tengan las astas más grandes, otros que sean capaces de saltar más alto, o tal vez una de las hembras que tenga un color más pardo que las otras. A nivel geográfico, las diferencias son aún más marcadas: los venados del norte del país son notablemente más grandes que su contraparte en el sureste del país, a pesar de pertenecer a la misma especie. Estas diferencias entre individuos son un componente importante de la biodiversidad que pasamos por alto al simplemente contar especies para cuantificar la riqueza biológica.
En otros niveles de organización existen otras manifestaciones de la biodiversidad. A nivel de las comunidades ecológicas, la composición de especies y las interacciones entre ellas varían considerablemente de un sitio a otro. Para cualquier persona resulta obvia la diferencia entre un desierto y una selva tropical en cuanto al tipo y número de especies de plantas y animales que se encuentran en estos biomas. Sin embargo, la estructura de esos conjuntos de organismos, por ejemplo en cuanto al número de individuos de cada especie y sus caractérísticas morfológicas, resulta menos evidente. Asimismo, la compleja red de interacciones entre los organismos es aún menos conspicua y sin embargo es una parte importante del fenómeno que llamamos biodiversidad.
El asteroide B 612 podrá ser muy pobre en especies, pero su único habitante puede jactarse de conocer y comprender en forma más profunda la biodiversidad de su hogar que cualquier ser humano del planeta Tierra. A través de sus actividades cotidianas, el Principito efectúa un programa eficiente de conservación y manejo de sus recursos. En efecto, al controlar el crecimiento de las plántulas de baobab, regar su preciada flor y deshollinar los tres volcanes del planetoide para evitar erupciones violentas, el habitante de B 612 usa, pero al mismo tiempo protege, la diversidad de este asteroide. Es más, al hacerlo, el pequeño y misterioso ser ama con pasión y se integra en cuerpo y alma a la riqueza biológica de su planeta, por magra que ésta pueda ser.
Desesperado por la codicia del hombre de negocios al contar estrellas, el Principito le espetó: “Es útil para mis volcanes y es útil para mi flor que yo los posea. Pero tú no eres útil a las estrellas”. Este concepto de posesión de la riqueza biológica a través del servicio mutuo es algo que debería hacernos pensar a la hora de diseñar proyectos de manejo de los recursos naturales. Asimismo, deberíamos ir más allá de los simples números, comprender más profundamente la relevancia y las implicaciones de algo tan complicado como lo es la biodiversidad y tratar de ver las miles de sutiles facetas que oculta el fenómeno de la diversidad de los organismos sobre nuestro planeta. Parafraseando al zorro filósofo que se encontró el Principito en la Tierra: la biodiversidad sólo se ve bien con el corazón. Su parte esencial es invisible para los ojos.
|
||||
|
Lecturas adicionales
Saint-Exupéry, A. 1943. Le Petit Prince. Existen muchas traducciones al español de este clásico que nos puede enseñar mucho sobre la biodiversidad y sobre la vida.
Los datos sobre el número de especies en México fueron obtenidos de: conabio. 1998. La diversidad biológica de México: estudio del país. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
|
||||
|
Héctor T. Arita
Instituto de Ecología,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Arita, Héctor T. (1999). La biodiversidad en el Asteroide B 612. Ciencias 53, enero-marzo, 40-42. [En línea] |
||||
 |
 |
|||
| personajes | ||||
|
Alfonso Nápoles Gándara
|
|
|||
|
Alberto Barajas
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
En 1930, el misterioso azar nos reunió a un joven profesor de matemáticas y a mí en un palacio. Ese año sería crucial en nuestras vidas, pero creo que ninguno de los dos lo presentía, aun cuando yo lo escuchaba atentamente disertar sobre las variaciones del trinomio de segundo grado.
La Universidad estaba estrenando la autonomía que conquistó en la lucha de 1929. En los mexicanos duraba todavía el estremecimiento provocado por las elecciones de 1928, en las que un general menospreció la Constitución y quiso reelegirse matando a sus dos contrincantes para asegurar su triunfo, aunque a su vez, después fue víctima, de una mano fanática instrumento de la justicia inmanente. El general Obregón murió exactamente el 17 de julio de 1928, día de mi cumpleaños.
Entré a la preparatoria lleno de expectativas, pues esperaba respuesta a muchas preguntas que me inquietaban: ¿en qué clase de país me había tocado nacer?, ¿qué sentido tiene la vida humana?, ¿qué cosa es la universidad y el amor y la cultura? Me obsesionaba, sobre todo, la pregunta que se han hecho los adolescentes de todos los tiempos: ¿qué rumbo le daré a mi vida?
Cada día estaba más desorientado y no podía dormir. Por otro lado juzgaba a mis profesores con frialdad implacable ya que me parecían menos interesantes que algunos, magníficos, que había tenido en la Secundaria 3. Pero existían dos excepciones: el joven Alfonso Nápoles Gándara y el viejo Erasmo Castellanos Quinto. Sí, con ellos las clases de matemáticas y de literatura universal eran muy gratas.
Como Kafka, fatigué desesperado los corredores del Palacio de San Ildefonso, y súbitamente empecé a escuchar voces que parecían venir de algún confín remoto para calmar mis dolores metafísicos. Una voz me dijo al oído: “No existen sino anhelos, Barajas. Lo demás no existe, Por los menos no existe vitalmente. La realidad de que habla la ciencia es realidad pensada; realidad viva sólo la tienen los objetos cuando en ellos se prende nuestro deseo o nuestra nostalgia. Tener las cosas no nos importa, importa aspirar a ellas o extrañarlas cuando ya se han ido. Parecemos los hombres una caravana que camina bajo el sol insoportable del desierto. Nos hacemos la ilusión de que somos mercaderes, pero lo que en verdad queremos es sentir sed. Sed de saber, sed de amar. Sed de gozar de sufrir; de vivir... de morir...”
Otra me dijo algo semejante con una metáfora: “Quiero doblar el arco de la vida hasta que forme un círculo. De mis manos saldrá entonces la flecha de la existencia que persigo. Correrá entre los bosques, saltará sobre un río... El aire, herido por su vuelo, irradiará: con luz de constelaciones que viene del infinito. ¿Podrá el arco doblarse, sin romperse, hasta formar un círculo? La flecha que saldrá de mi arco ¿llegará a su destino?” El mundo empezaba a aclarárseme.
El profesor Nápoles caminaba muy aprisa y con la mirada hacia adelante. Siempre iba muy derecho pues tenía una de esas espaldas sin curvatura que sólo se ven en el Colegio Militar. Fuera de clase parecía no reconocernos, en cambio, en clase... tampoco. Sus exposiciones eran muy claras, ya que calculaba muy bien las dosis de conocimiento que podíamos absorber sin mayor esfuerzo. Escribía en el pizarrón lo que era necesario y suficiente, y no nos abrumaba con dictados inútiles. Su voz era muy clara y se escuchaba perfectamente hasta la última fila.
Un día empezamos a notarlo nervioso y preocupado, entonces nos anunció que el curso tendría que acortarse y el examen final sería en agosto. Por los periódicos supimos la causa de su inquietud: le acababan de conceder la beca Guggenheim para hacer estudios superiores en el Instituto Tecnológico de Massachusetts. Era el primer matemático mexicano que obtenía tal honor. A partir de ese momento, nuestro respeto por él dio un salto cuántico.
Al darnos las calificaciones del examen comentó brevemente mi trabajo. Su demostración es interesante, me dijo. Tiene diez. Esa vez fue el único que puso.
Para Nápoles fue seguramente muy angustioso el contacto brusco con el mundo de la ciencia internacional. En aquella época había una gran distancia entre la preparación matemática que se podía obtener en México y la que ofrecían las grandes universidades extranjeras. Sólo el que ha estado en ellas puede apreciar el esfuerzo heroico de Alfonso Nápoles para acreditar catorce cursos semestrales de matemáticas superiores de categoría A con la máxima calificación de H, aprobado con honor, en once de ellos. Si consideramos que esto ocurrió en un lapso de año y medio, su actuación fue muy notable.
Regresó a México en 1932 con un tesoro de conocimientos que ha compartido con sus discípulos y sin los cuales no hubiera sido posible la colaboración con grandes científicos extranjeros. Cuando yo discutía con Einstein una teoría rival a la suya, la de Birkhoff, pensaba que esa conversación hubiera sido imposible sin el curso de Cálculo Tensorial que tomé con don Alfonso. Al pensar en su vida recuerdo inevitablemente las palabras que también escuché en 1930, mi año terrible, y que ya he mencionado en alguna otra ocasión: “Hay hombres que sólo entran al combate cuando el Rey está mirando. O son como Aristo, aquel filósofo tan elegante que sólo filosofaba cuando sus amigos lo llevaban en una litera lujosa. Hay otros, en cambio, que trabajan siempre que se necesita y en las condiciones menos favorables; dispuestos todo el tiempo a cumplir con sus deberes y sus ideales”.
La biografía de Nápoles nos muestra su devoción por las matemáticas y por la Universidad. Esta prodigiosa Universidad de México que tanto me deconcertó al principio y que al fin me reveló cuál era mi destino, mi dharma. Si en un momento de crisis no hubiera tenido un profesor de matemáticas tan distinguido, probablemente habría seguido otra carrera.
Alfonso Nápoles Gándara nació en Cuernavaca el 14 de octubre de 1897. Allí estudio la primaria; después estuvo en la Escuela Nacional Preparatoria y en la Escuela Nacional de Ingenieros, donde por su talento y originalidad llamó la atención del extraordinario Sotero Prieto.
Cuando Sotero murió, asumío el liderazgo del movimiento matemático en México. Más adelante, formó parte del grupo fundador de la actual Facultad de Ciencias, donde dirigió el Departamento de Matemáticas de 1939 a 1965. En 1942 se fundó el Instituto de Matemáticas, del que fue su primer director en noviembre de 1942, inició en Saltillo la serie de congresos de matemáticas que se han venido realizando con tanto éxito a lo largo de muchos años. Como consecuencia del primer congreso se creó la Sociedad Matemática Mexicana, la cual presidió hasta 1961. Desde entonces es Presidente Honorario Vitalicio.
Está casado con la señora Guadalupe Salazar y su hijo es el arquitecto Alfonso Nápoles Salazar, es hombre muy inteligente y muy creativo, quien además es distinguido y respetado en su profesión.
En fin, como quería el poeta, don Alfonso supo empuñar el arco con valentía y firmeza para lograr que la flecha llegara a su destino.
Septiembre de 1991
|
||||
|
Alberto Barajas
Instituto de Matemáticas,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Barajas Celis, Alberto.(1999). Alfonso Nápoles Gándara. Ciencias 53, enero-marzo, 28-30. [En línea]
|
||||
|
|
||||
 |
 |
|||
| de lo solube y lo insoluble |
||||
|
Reflexiones en torno al binomio
ciencia-sociedad
|
|
|||
|
Antonio R. Cabral
conoce más del autor
|
||||
| HTML ↓ | ← Regresar al índice ⁄ artículo siguiente → | |||
|
Ya es un lugar común decir que, en los últimos cincuenta años, la ciencia ha influido a la sociedad más que en toda su historia anterior. La biología molecular, en otro tiempo considerada como tema de novelas, es ahora uno de los motores más eficaces de la ciencia moderna. La ingeniería genética, la clonación de ovejas, los análisis de ácidos nucleicos depositados en vestidos azules son noticias de ocho columnas. Por todos lados se nota el impacto de la ciencia en la colectividad; esto es tan obvio que parece ya no haber nada más qué decir. La ciencia, y su sucedánea, la tecnología, son parte de prácticamente todas las actividades humanas, ya que de una u otra manera están inmersas no sólo en las casas que habitamos, en los alimentos que comemos y en los transportes que abordamos, sino también en los artículos electrónicos que usamos para trabajar o para no aburrirnos. Podría decirse que en mayor o menor grado, nadie escapa del arrope de la ciencia. Por ello, ahora que al parecer el planeta se ha librado de la destrucción nuclear, la sociedad presta atención a otros peligros atribuidos a la ciencia como la clonación de humanos, la creación ad libitum de virus tan temidos como letales y la manufactura de órganos a destajo. Los científicos podrían a su vez ser vistos como seres misteriosos que, metidos en sus laboratorios, maquinan ideas perversas de cómo dañar a sus semejantes sin dejar la menor huella; puede que esto no sea totalmente falso: no sería difícil que alguien con conocimientos profundos sobre la vida misma y que al mismo tiempo padezca debilidades humanas, quiera aplicar esos conocimientos para hacer el mal. Tampoco debe dudarse de la existencia de científicos que prefieran seguir otros caminos que los señalados por la ética científica, que no es otra que la ética cotidiana, y utilicen su experiencia para beneficio propio sin que medie el más mínimo interés en buscar la verdad por la verdad misma. En un mundo tan competitivo como el que ha tocado vivir no deben extrañar esos desvíos: los científicos, como los empresarios, los políticos, los choferes, los taxistas, los bibliotecarios, los sacerdotes y el público en general, nunca han pertenecido a la especie que tiene un nimbo sobre sus cabezas.
Con todo y las innovaciones tecnológicas, las asimetrías sociales están más presentes que nunca: el paludismo sigue matando a varios miles de personas al año en países africanos a pesar de que, en 1902, Sir Ronald Ross recibió el Premio Nobel de Medicina precisamente por sus descubrimientos sobre el ciclo vital del parásito causante de esa enfermedad. Igualmente, en las sociedades avanzadas una gran proporción de las riquezas nacionales se gastan en salud y recreación, pero en el mundo subdesarrollado las hambrunas y las guerras matan a millones de personas, la desnutrición y las enfermedades parasitarias son muy frecuentes y ni siquiera se pueden satisfacer las necesidades más básicas como la comida y la habitación. Desde luego que a la ciencia le gustaría decir que ya tiene la solución para la pobreza y que pronto todos los habitantes de la Tierra tendrán alimentos sobre sus mesas tres veces al día. Todos sabemos que estos problemas, por ser del dominio de los gobiernos y de su política económica, rebasan los límites de la ciencia, y aun los de la economía, la cual, dicen sus practicantes, es parte de ella. Pero no por esto, la sociedad debe sentirse defraudada; al contrario, debe estar segura de que la ciencia, al hacerse preguntas que pertenecen al mundo real, tarde o temprano, encontrará respuestas que tengan una aplicación práctica. En la mayoría de los países la investigación científica está patrocinada principalmente por agencias gubernamentales, ya que la participación de la industria privada en los países subdesarrollados, con algunas excepciones, es magra. La sociedad analiza los logros de los hombres de ciencia mediante comisiones evaluadoras de la actividad científica y confía en que el dinero destinado para ellos es bien utilizado. En los países desarrollados, el gasto asignado a la investigación científica se escatima cada vez menos, pues los gobernantes saben que esto trae consigo bienestar y riqueza para los ciudadanos y con ello, hay que decirlo, la posibilidad de reelegirse; pero en los países menos favorecidos, la sociedad aún no se ha dado cuenta de que la ciencia es un vehículo seguro para salir del subdesarrollo y no al revés. Según lo apuntó nuestro admirado Voltaire, esto no sorprende: es difícil que la razón prevalezca ahí donde domina la superstición. Mientras en Estados Unidos el número de científicos por cada diez mil habitantes es de treinta y cinco, en México es de uno. No debemos imitar el camino andado por los estadunidenses porque sea el mejor modelo, sino porque, con todo y sus limitaciones, la ciencia ofrece alternativas razonables, y para, como dice el doctor Ignacio Chávez, “no pasarnos la vida rezando las verdades y los errores que nos legaron otros”.
El quid no es la falta de creatividad, ya que los científicos que habitan el Tercer Mundo no son cortos de ideas, sino que padecen la enfermedad que el biólogo molecular inglés avecindado en California, Sydney Brenner, reciente y atinadamente bautizó como mdd, que en nuestro idioma bien podría traducirse con puntualidad como “Millones de Dólares”, pero que en el de él es “Money Deficiency Disease”. A nadie conviene padecer la enfermedad de Brenner; cuando ésta ataca sólo a una persona, las secuelas no son muy graves porque no afectan directamente a la sociedad. En cambio, cuando la ciencia y la tecnología de un país enferman de mdd, los síntomas rebasan al paciente y sus repercusiones son incalculables. Dinero y bienestar están íntimamente ligados, no en vano el doctor Amartya Sen recibió este año el Premio Nobel de Economía por sus estudios sobre la calidad de vida, concepto éste que pocos entienden y que nadie sabe definir, pero que pocos pueden rebatir. El impacto de la ciencia en la colectividad, ya quedó dicho, es indiscutible. Desde hace algunos años diversos científicos estadunidenses intentan convencer a sus compatriotas para que dejen entrar la ciencia en sus modelos de pensamiento y, consecuentemente, en su acción. Ya se verán los efectos. En nuestro país hay esfuerzos en el mismo sentido, aunque, seguramente, son insuficientes. La afrenta es doble: producir a pesar de la mdd e incrementar la cultura científica de un país en donde la ciencia no es un valor.
|
||||
|
Antonio R. Cabral
Instituto Nacional de la Nutrición “Salvador Zubirán”.
_______________________________________________________________ como citar este artículo →
Cabral R., Antonio.(1999). Reflexiones en torno al binomio ciencia-sociedad. Ciencias 53, enero-marzo, 14-15. [En línea]
|
||||
|
|
||||
 |
 |
||||
|
|
|||||
|
Las metapoblaciones en la naturaleza, ¿realidad o fantasía?
|
|||||
|
María Teresa Valverde Valdés
conoce más del autor
|
|||||
|
|
← Regresar al índice ⁄ artículo siguiente → | ||||
|
Cuando estudiamos la dinámica de una población ecológica (entendida ésta como un grupo de organismos de la misma especie que coexiste en un espacio común) generalmente nos centramos en el análisis de los parámetros demográficos básicos, a saber, las tasas de natalidad, de mortalidad y de migración. Sin embargo, es muy común que este último término sea ignorado (i.e., la migración) y que se parta de la base de que los únicos procesos que determinan los cambios numéricos de una población son los nacimientos y las muertes de los individuos. La teoría metapoblacional se encuentra fincada en el hecho de que la migración entre las poblaciones de una misma especie no siempre es despreciable, sino que por el contrario, con frecuencia es de gran importancia, pues determina la dinámica espacial de los conjuntos de poblaciones de una misma especie a través del tiempo. Esta idea nos lleva directamente a la definición del concepto de metapoblación: las diferentes poblaciones de una especie no se encuentran aisladas en la naturaleza, sino que mantienen cierto contacto entre ellas a través de la migración de individuos; a estos conjuntos o grupos de poblaciones de una misma especie, interconectados por la migración de individuos entre ellas, se les conoce como metapoblaciones.
Una de las ideas implícitas en el concepto metapoblacional es la suposición de que las poblaciones que forman una metapoblación (llamadas “poblaciones locales”) no son eternas, sino que pueden extinguirse por diversas causas. De la misma forma, algunos de los sitios o “parches” del ambiente susceptibles de ser colonizados por la especie pueden encontrarse vacíos y la migración de individuos hacia ellos puede dar lugar a la formación de nuevas poblaciones. De esta manera, una metapoblación puede concebirse como un sistema dinámico en el que constantemente ocurren la formación y la extinción de poblaciones locales. Así, de manera análoga al estudio de una población, cuya dinámica se explica con base en las tasas de natalidad y mortalidad, el estudio de la dinámica de una metapoblación se basa en el conocimiento de las tasas de colonización y extinción de poblaciones locales en un contexto regional, y en el análisis de las condiciones que permiten un equilibrio entre ambos procesos. Dado que las especies que forman metapoblaciones se encuentran sujetas a altos riesgos de extinción de sus poblaciones locales, su permanencia a nivel regional depende del balance entre colonizaciones y extinciones. Surge el concepto metapoblacional Al analizar la historia del concepto metapoblacional, aparentemente nuevo en ecología, causa sorpresa encontrarse con el hecho de que dicho concepto data de finales de la década de los sesentas. De hecho, la idea básica de lo que en la actualidad se conoce como una metapoblación se encuentra ya en textos ecológicos clásicos, como el de Andrewartha y Birch, incluso anterior a esas fechas. Sin embargo, no es sino hasta finales de los sesentas que Richard Levins, un entomólogo interesado en el control de plagas, “bautiza” a las metapoblaciones y formaliza el concepto mediante, de la propo sición del primer modelo metapoblacional, que posteriormente daría pie al desarrollo de lo que ahora conocemos como teoría metapoblacional. Es interesante analizar el contexto histórico en el que Levins propone, por vez primera, la existencia de metapoblaciones en la naturaleza: desde los años treintas y hasta finales de la década de los cincuentas se dio una controversia importante y de consecuencias muy fructíferas entre los ecólogos poblacionales, que tenía que ver con la discusión sobre los factores que regulan el tamaño de las poblaciones en la naturaleza. En esa época existían dos escuelas principales de pensamiento: a) una que sostenía que los factores reguladores del tamaño poblacional eran de naturaleza biótica (depredación, parasitismo, competencia) y que por lo tanto se manifestaban con mayor intensidad en poblaciones con densidades altas, y b) otra que planteaba que eran principalmente los factores abióticos (tormentas, perturbaciones, heladas, etcétera) y, por lo tanto, independientes de la densidad poblacional, los que mantenían a las poblaciones alrededor de ciertos valores de densidad. Una tercera alternativa que recibió poca atención en ese momento planteaba que los mecanismos de regulación poblacional no son tan eficientes como se suponía, y que si se seguía a una población por un tiempo suficientemente prolongado se observaría que, a la larga, tendería a la extinción. Entonces, la permanencia a nivel regional de especies con este tipo de comportamiento numérico debía estar determinada por un balance entre la extinción de poblaciones locales y la formación de poblaciones nuevas (a través de la migración) al interior de un “ensamble” de poblaciones locales efímeras.
No es coincidencia que haya sido precisamente Levins, un entomólogo especializado en el control de plagas, el que desarrolló esta idea y la plasmó en el modelo metapoblacional básico (que se presenta más adelante), pues si analizamos el desarrollo regional de cualquier plaga nos daremos cuenta de que ésta se comporta como metapoblaciones: las plagas no tienen mecanismos eficientes de control poblacional, sino que llegan a un lugar, consumen todos los recursos disponibles y posteriormente desaparecen. El hecho de que a pesar de estas extinciones muchas plagas sean tan difíciles de erradicar es un reflejo de que su permanencia está dada por un balance entre las extinciones y la formación de nuevas poblaciones a través de la colonización de sitios suceptibles de ser explotados. Algunos de los grandes aciertos de Levins al proponer el concepto metapoblacional son, por un lado, la descripción de esta dinámica mediante del planteamiento de un modelo matemático sencillo y, por otro lado, la sugerencia implícita de que posiblemente muchas otras especies, además de las plagas, presenten este tipo de dinámicas espacio-temporales, detectables únicamente por encima del nivel de análisis estrictamente poblacional. La propuesta de Levins El modelo de Levins considera especies que muestran patrones de distribución espaciales a manera de “parches” discretos y bien definidos en los que la especie puede vivir inmersa en una “matriz” de hábitat formada por ambientes no colonizables. El modelo considera únicamente los parches en los que la especie puede establecerse (por ejemplo, los árboles hospederos de una planta epífita en un bosque, o las charcas de agua que se forman en las praderas y que son colonizadas por ciertas especies de ranas), y describe el estado de la metapoblación en un momento dado en términos de la proporción de estos parches que se encuentran realmente ocupados por la especie y la proporción de parches que se encuentran vacantes. El modelo estima el cambio en la proporción de parches ocupados a través del tiempo, tomando en cuenta la tasa de colonización de parches (formación de poblaciones locales nuevas) y la tasa a la que las poblaciones locales se extinguen (ver recuadro).
Es importante hacer notar que el modelo metapoblacional de Levins es prácticamente idéntico en su forma a algunos de los modelos desarrollados en epidemiología para describir la dinámica de las infecciones por parásitos. Esto es reflejo de que, en ambos casos, se concibe a una especie (el parásito, en el caso de los modelos epidemiológicos) que va “infectando” ciertos ambientes colonizables mientras es erradicada de otros, generando la dinámica de colonización–extinción que caracteriza a ambos sistemas.
El modelo metapoblacional de Levins está basado en varios supuestos fundamentales: 1) la extinción sucede y, por lo tanto, la permanencia de una especie en una región debe depender de la colonización; 2) todos los parches disponibles tienen la misma probabilidad de ser colonizados, es decir, no hay un efecto significativo de la distribución espacial o del nivel de aislamiento de los parches; 3) la tasa de colonización es proporcional a la fracción de parches ocupados; 4) todas las poblaciones tienen la misma probabilidad de extinguirse, pues no hay un efecto del tamaño, la longevidad o la dinámica interna de las poblaciones locales, y 5) la probabilidad de extinción de cada población es independiente, ya que no ocurren extinciones correlacionadas.
Es claro que estos supuestos no se cumplen en las metapoblaciones que encontramos en la naturaleza, pero también es cierto que la mayoría de ellos pueden relajarse sin necesidad de hacer modificaciones conceptuales significativas al modelo. En realidad, utilizar un modelo metapoblacional con estas características no es muy distinto de la forma en la que se utiliza el concepto de “población” en biología poblacional: ninguna población real satisface cabalmente los criterios de ser una entidad panmíctica (en donde la reproducción puede darse entre todos los individuos que la conforman) y cerrada (sin migración), y a pesar de todo el concepto poblacional es útil e ilustrativo para abordar el estudio de una parte importante de la naturaleza viva. De la misma forma, el concepto de metapoblación ofrece una concepción particular sobre la estructura espacial de las especies en la naturaleza, que nos habla de la existencia de poblaciones locales discretas, conectadas a través de la migración de individuos entre ellas. Esta concepción, en sí misma, es de gran valor, pues considera una escala espacial (regional) que no había sido abordada por la ecología poblacional; enfatiza procesos que se dan en una escala temporal y espacial que no siempre es obvia para el ojo humano, y, además, constituye un salto desde la ecología descriptiva, que señala la existencia de patrones en la naturaleza, hasta la ecología analítica, que se preocupa por describir los procesos que dan lugar a dichos patrones. Extinción y tipos de metapoblaciones El concepto metapoblacional y, consecuentemente, toda la teoría que se desprende de él, dependen de la idea de que la extinción de las poblaciones locales ocurre (y de ahí la importancia de la migración para permitir la recolonización). En efecto, en la literatura se encuentran reportados un gran número de casos en los que se ha documentado la extinción de poblaciones locales de diversas especies de animales y plantas, lo cual sugiere que las dinámicas de colonización–extinción podrían ser importantes en algunos sistemas naturales. Las causas que han provocado estas extinciones pueden ser de tipo estocástico (es decir, debidas al azar) o de naturaleza determinística. Las extinciones estocásticas tienen que ver con tres tipos fundamentales de procesos azarosos: 1) aquellos que ocurren a nivel ambiental (tales como las variaciones en las condiciones climáticas, las heladas severas, las tormentas, los deslaves, los ciclones, los incendios, etcétera); 2) los que toman lugar a nivel de la dinámica interna de las poblaciones locales (i.e., “estocasticidad demográfica”, dada por variaciones aleatorias en el orden y magnitud de los nacimientos y muertes de individuos al interior de una población), y 3) los que tienen que ver con las características genéticas de las poblaciones (i.e., “estocasticidad genética”, en donde los procesos de deriva génica pueden provocar pérdida de variabilidad genética y depresión endogámica y llevar a una población a la extinción).
Las extinciones determinísticas (es decir, ocasionadas por procesos no azarosos) también parecen ser relativamente frecuentes. Éstas ocurren como producto de cambios direccionales en las condiciones del medio que poco a poco provocan que el parche ocupado por una especie se transforme en un sitio inhabitable. Esto ocurre, por ejemplo, en los hábitats de carácter efímero o temporal, tales como una fruta que cae al suelo en un bosque y se convierte en un parche susceptible de ser colonizado por larvas de insectos, hongos, etcétera, pero que a la larga desaparecerá. Otro tipo de extinciones determinísticas son las que se dan en especies que ocupan hábitats sucesionales tempranos dentro de algunas comunidades. Por ejemplo, muchas especies de plantas colonizan los claros que se forman en los bosques cuando ocurre una pequeña perturbación, como la caída de un árbol o una rama grande; estos claros se convierten en parches que pueden ser colonizados por especies de plantas que requieren de este tipo de condiciones ambientales para establecerse, sin embargo, al irse cerrando el claro a causa del crecimiento de la vegetación adyacente (proceso conocido como sucesión secundaria), el parche se convierte poco a poco en un sitio inhabitable y la población local que lo haya ocupado estará destinada a la extinción. Basada en la relevancia a nivel regional de los procesos de colonización–extinción para diferentes especies, Harrison propone la existencia de cuatro tipos de metapoblaciones: a) Metapoblaciones tipo isla-continente. Son aquellas en las que existe una población grande que funciona como la fuente principal de colonizadores de los parches pequeños cercanos a ella. En este tipo de metapoblaciones las extinciones se dan únicamente en las poblaciones pequeñas (las “islas”), mientras que la población grande (el “continente”) es prácticamente inmune a la extinción. Consecuentemente, sí existe una dinámica de colonización–extinción, pero ésta no determina la permanencia de la especie a nivel regional. Este tipo de metapoblaciones se han observado en algunas especies de mariposas. b) Metapoblaciones tipo mosaico. Son aquellas en las que la especie presenta, en efecto, una distribución a manera de parches discretos, pero entre los cuales la migración de individuos es tan generalizada que no se puede considerar a cada parche como una población local relativamente independiente. El conjunto de parches forma, en realidad, una entidad demográfica única e integrada y el alcance tan generalizado de la migración no permite la extinción. Algunas especies sucesionales, como las que se describieron en el párrafo anterior, parecen tener este tipo de estructura metapoblacional. c) Metapoblaciones no equilibradas. Muchas especies que originalmente no presentaban una estructura metapoblacional se han visto recluidas, en las últimas décadas, a pequeños fragmentos de hábitat natural rodeados de ambientes alterados, como los campos de cultivo o los pastizales de uso ganadero. Estas especies presentan ahora muchas de las características propias de las metapoblaciones, tales como la existencia de parches colonizables rodeados de una matriz de hábitat no colonizable y la alta ocurrencia de extinciones locales. Sin embargo, la re-colonización de los parches vacantes se ve limitada por la baja capacidad de migración de los individuos y, por lo tanto, la tasa de extinción excede a la tasa de colonización. Así, este tipo de metapoblaciones se presentan en el contexto de una disminución generalizada en la abundancia de muchas especies, determinada por el deterioro paulatino del hábitat. Muchas especies de mamíferos grandes, comoalgunos felinos y osos, presentan este tipo de metapoblaciones. d) Metapoblaciones clásicas. Son aquellas que se acercan en mayor medida a la idea de metapoblación propuesta en el modelo original de Levins. En estos sistemas existe un balance real entre colonizaciones y extinciones, que es lo que determina la permanencia de la especie a nivel regional. Se han documentado muy pocos casos que cumplan estrictamente con estas características; uno de los más notables corresponde a un sistema de charcas que son colonizadas por una especie de rana y en donde son frecuentes tanto la extinción de poblaciones locales como la formación de poblaciones nuevas. Metapoblaciones y conservación El concepto metapoblacional propuesto por Levins no tuvo mucho eco en la comunidad de ecólogos en su momento, sino que pasó prácticamente inadvertido hasta ya bien entrada la década de los ochentas, cuando se retomó y se comenzó a elaborar poco a poco para dar paso a la actual teoría metapoblacional. Los temas de interés entre los ecólogos poblacionales de los sesentas eran hasta cierto punto distintos de los que ahora constituyen el foco de atención de los ecólogos contemporáneos, y la visión de Levins no adquirió la generalidad necesaria como para llamar la atención en un círculo amplio de investigadores. En contraste, uno de los temas de vanguardia de la ecología contemporánea es precisamente la dinámica y la estructura espacial de los sistemas ecológicos. Es quizá por esta razón que la teoría metapoblacional parece haber encontrado su nicho natural en la ecología moderna, al ofrecer una visión particular de la dinámica espacial entre grupos de organismos de la misma especie. La teoría metapoblacional actual va incluso más allá del nivel exclusivamente uni–específico y aborda el área de las interacciones ecológicas, como la competencia y la depredación, planteando la forma en la que ciertos sistemas de pares de especies (competidoras y depredador–presa) pueden coexistir en equilibrio gracias a la existencia de estructuras metapoblacionales. Sin embargo, más que en el ámbito de la ecología tradicional, la teoría metapoblacional ha tenido un impacto importante en el área de biología de la conservación. Quizá el auge de esta disciplina en tiempos recientes es otra de las razones que han determinado el resurgimiento de la teoría metapoblacional. Un hecho que bien da cuenta de esta asociación entre la biología de la conservación y la teoría metapoblacional es que esta última aborda formalmente el tema de la extinción; además, al analizar la dinámica espacial de especies de interés particular, ofrece una serie de criterios ecológicos sólidos para apoyar el diseño de reservas, corredores ecológicos, áreas protegidas y demás. Respecto de este último punto, es interesante notar que durante la década de los años setenta el paradigma de la biología de la conservación, sobre todo en lo referente al diseño de reservas, fue la teoría de biogeografía de islas, planteada por MacArthur y Wilson en 1967, mientras que ahora es la teoría metapoblacional la que generalmente se invoca para ofrecer criterios ecológicos en la toma de decisiones sobre áreas protegidas. Para entender las causas que han determinado este cambio de paradigmas es necesario analizar la contribución particular de cada una de estas dos teorías a la biología de la conservación.
Uno de los aspectos más importantes de la teoría de biogeografía de islas que posteriormente fue retomado por la biología de la conservación, fue la idea de que existe una relación entre el área ocupada por una cierta comunidad ecológica y el número de especies que puede mantener. Además, MacArthur y Wilson analizaron la relación entre la distancia a la que una “isla” (un fragmento aislado de hábitat colonizable) se encuentra del “continente” (es decir, de la fuente de colonizadores), y la tasa de llegada y desaparición de especies de esa isla, sugiriendo que el número de especies presentes en ella llegaría a un cierto equilibrio, dado precisamente por el área de la isla. Estas ideas parecían enfatizar el uso de determinadas estrategias, relacionadas con el tamaño, forma y disposición de las áreas protegidas, destinadas a maximizar el número de especies que albergarían. A partir de esto, los conservacionistas comenzaron a manejar una serie de “reglas” para el diseño de reservas que, incluso, a la fecha, se asocian con la teoría de biogeografía de islas. Algunas de estas “reglas” son, por ejemplo, la noción de que un área grande puede albergar una mayor diversidad que un área pequeña, o que áreas contiguas conectadas por corredores presentan más especies que áreas aisladas de tamaño equivalente. Nótese que a pesar de que existe una cierta similitud en el contexto en que se plantean las teorías metapoblacional y de biogeografía de islas (pues ambas visualizan el ambiente como un conjunto de parches colonizables rodeados de una matriz de ambiente no colonizable, además de que enfatizan el análisis de las tasas de colonización y extinción), éstas abordan aspectos muy diferentes de la ecología. La teoría de biogeografía de islas tiene que ver con la dinámica de conjuntos de especies y con el análisis de los factores que determinan la riqueza específica al interior de los parches colonizables presentes en una cierta área. La teoría metapoblacional, por su parte, analiza la dinámica espacial de una especie en particular y estudia las condiciones que afectan los cambios en el número de parches ocupados por dicha especie. Dicho en otras palabras y de manera un tanto simplista, la teoría metapoblacional forma parte de la ecología de poblaciones, mientras que la teoría de biogeografía de islas es claramente parte del ámbito de la ecología de comunidades. En resumen, estas dos teorías buscan respuestas a preguntas esencialmente diferentes y, por lo tanto, su contribución a la biología de la conservación, sobre todo en lo relacionado con el diseño de reservas, no es ni siquiera comparable, pues abordan aspectos distintos de la ecología. La teoría metapoblacional ofrece elementos para establecer criterios útiles para la creación de reservas, pero no desde la óptica de la diversidad de la comunidad en su conjunto, sino de la permanencia de especies particulares a nivel regional a través del conocimiento de la dinámica espacial de sus poblaciones. El cambio de paradigmas al que se ha hecho referencia con anterioridad parece tener varias causas. Una de ellas está relacionada con el hecho de que, como era de esperarse, no se han encontrado suficientes evidencias empíricas que apoyen las predicciones derivadas de la teoría de biogeografía de islas. Por otro lado, la biología de la conservación parece estar dando un giro del enfoque comunitario al poblacional en los últimos años (en el cual puede incluirse el reciente interés por los aspectos genéticos de las poblaciones), lo cual ha favorecido el auge de la teoría metapoblacional. Además, la adquisición de un enfoque metapoblacional ha permitido analizar los paisajes naturales como “redes” o sistemas de reservas, y no tanto como reservas aisladas, haciendo énfasis precisamente en las dinámicas espaciales de los sistemas naturales, que constituyen una de las preocupaciones fundamentales de la ecología moderna. ¿Por qué este enfoque? La breve exposición hecha en los párrafos anteriores sobre la importancia de la teoría metapoblacional en ecología de la conservación no es exhaustiva, ni pretende serlo; sin embargo, sí señala algunos de los aspectos de mayor relevancia que permiten explicar su utilidad desde el punto de vista teórico y conceptual. Ahora, es necesario tomar en cuenta que las metapoblaciones, como entidades reales en la naturaleza, pueden ser muy difíciles de delimitar, estudiar, o sencillamente observar, pues no nada más se presentan a escalas espaciales generalmente difíciles de manejar, sino que, además, prácticamente nunca cumplen con los supuestos del modelo metapoblacional original. Por otro lado, el movimiento de individuos entre poblaciones de una misma especie puede ser extremadamente difícil de cuantificar y describir, lo cual es una limitación muy seria si se toma en cuenta que la migración es una de las condiciones básicas del concepto metapoblacional. Todo esto ha llevado a ciertos autores, como Susan Harrison, a cuestionar la utilidad del concepto en su forma original un tanto rígida, a causa de la falta de evidencia empírica que lo apoye. De ahí el título del presente artículo, que hace referencia precisamente a esta crítica. Para responder en alguna medida a la crítica de Harrison, me gustaría retomar la relación entre los conceptos teóricos de población y de metapoblación, con relación a que ninguno de ellos refleja realmente ningún sistema natural en el sentido estricto de la palabra y que, a pesar de todo, ambos nos han sido útiles para describir y estudiar una parte importante de la naturaleza viva. Esta analogía, además, enfatiza uno de los puntos que me parece importante comentar en el presente artículo: el hecho de que los conceptos ecológicos no tienen que reflejar la realidad al cien por ciento para sernos útiles o para que los escojamos como una representación válida de algún aspecto de la realidad. Muchos de estos conceptos representan modelos de la realidad y, como tales, no la reflejan (ni pretenden hacerlo) en toda su magnitud y complejidad, sino que constituyen versiones simplificadas de los sistemas, fenómenos o procesos que nos interesa abordar; el único objetivo de esta simplificación es ofrecer una mayor accesibilidad al objeto de estudio en cuestión. En el caso del concepto metapoblacional, lo que es importante rescatar es esta concepción de que los grupos de poblaciones locales discretas de una especie, conectados a través de la migración de individuos entre ellas, pueden ser analizados a una escala espacial regional, por encima de la escala poblacional. Quizá no sea tan importante determinar con exactitud cuánta migración hay entre las poblaciones de algún sistema que nos interese estudiar, ni qué alcance espacial tiene esta migración, sino más bien decidir si al utilizar una visión metapoblacional vamos a alcanzar una mejor comprensión de ciertos procesos ecológicos que pasaríamos por alto si nos limitaramos a una visión exclusivamente poblacional. Desde un punto de vista poblacional o demográfico, nos podríamos formular la siguiente pregunta: ¿es válida la suposición de que las poblaciones son sistemas cerrados en donde el efecto de la migración puede ignorarse? Siempre que la respuesta a esta pregunta sea no, entonces podemos estar seguros de que el uso de una visión metapoblacional enriquecerá sustancialmente nuestra comprensión del sistema bajo estudio. |
|||||
|
Referencias bibliográficas
Andrewartha, H. y L. Birch. 1954. The distribution anda abundance of animals., Chicago, University of Chicago Press.
Hanski, I. y D. Simberloff. 1997. “The metapopulation approach, its history, conceptual domain, and application to conservation”, en Hanski, I. y M. E. Gilpin, eds., Metapopulation biology. Nueva York, Academic Press Inc., pp. 5-26. Harrison, S. 1991. Local extinction in a metapopulation context: an empirical evaluation. Biological Journal of the Linnean Society, 42 pp. 73-88. Levins, R. 1969. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bulletin of the Entomological Society of America, 15, pp. 237-240. Levins, R. 1970. Extinction. Lectures on Mathematics in the Life Sciences, 2, pp. 75-107. MacArthur, R y E. Wilson. 1967. The theory of island biogeography., Princeton, Princeton University Press. |
|||||
|
María Teresa Valverde Valdés
Facultad de Ciencias,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo →
Valverde Valdés, Maria Teresa. (1999). Las metapoblaciones en la naturaleza, ¿realidad o fantasía? Ciencias 53, enero-marzo, 56-63. [En línea] |
|||||
| ← Regresar al índice ⁄ artículo siguiente → |
 |
 |
||
|
|
|||
|
T. S. Kuhn y la “naturalización” de la filosofía de la ciencia
|
|||
| Ana Rosa Pérez Ransanz
conoce más del autor
|
|||
|
|
← Regresar al índice ⁄ artículo siguiente → | ||
|
Hoy día se acepta ampliamente que la ciencia es un fenómeno cultural complejo, cuyo desarrollo depende de múltiples factores: biológicos, psicológicos, sociales, económicos, técnicos, legales, políticos, ideológicos, etcétera. De aquí que la empresa científica se preste a ser analizada desde perspectivas teóricas muy diversas y en función de distintos objetivos e intereses. Tal parecería, entonces, que la reflexión sobre la ciencia no puede ser tarea de una sola disciplina.
Consideremos, por ejemplo, el hecho de que las teorías científicas son producto de una actividad humana colectiva, la cual se desarrolla en determinadas condiciones y estructuras sociales. Si esto es así, el estudio de las instituciones y de las comunidades donde se lleva a cabo esa actividad, o del impacto que tienen sus resultados en nuestras formas de vida, cubre aspectos importantes de la ciencia en tanto que fenómeno social. Por otra parte, la ciencia, como conjunto de prácticas y resultados, está sujeta a un proceso de cambio y evolución. Se trata de prácticas y productos que se generan y desarrollan, modifican o abandonan, a través del tiempo. De aquí que la comprensión de la dinámica científica requiera la información que generan los estudios históricos de las diversas disciplinas. También encontramos que los estudios psicológicos y neurofisiológicos de procesos cognitivos como la percepción, el aprendizaje o la invención, contribuyen a esclarecer ciertas condiciones de la producción y transmisión de conocimientos. Ahora bien, desde la perspectiva de la ciencia como una actividad de resolución de problemas, es decir, como una actividad dirigida al logro de objetivos específicos, cae también bajo el dominio de la teoría de las decisiones. Y en tanto que sistema de procesamiento de información, la actividad científica es analizada por quienes se ocupan del diseño y funcionamiento de dichos sistemas, los teóricos de la inteligencia artificial, y en general por quienes trabajan en el campo de la psicología computacional. Este somero e incompleto recuento permite entrever la amplia gama de investigación científica que hoy día se hace sobre la ciencia misma. Además, hay que destacar el hecho de que las diversas ciencias que se ocupan de la ciencia, las llamadas “metaciencias”, han tenido un desarrollo sin precedentes en las últimas dos décadas. Sin embargo, esta proliferación de estudios metacientíficos ha dado lugar a una intrincada controversia. Si bien actualmente domina la tendencia a considerar que la ciencia debe ser objeto de estudio de un gran programa interdisciplinario de investigación —quizá uno de los más complejos de la ciencia contemporánea—, por otra parte impera un considerable desacuerdo en cuanto al orden de importancia y las relaciones que guardan entre sí los distintos estudios sobre la ciencia. Incluso se destacan algunos grupos que pretenden que su perspectiva teórica tiene un carácter fundamental o privilegiado. A modo de ejemplo consideremos una posición radical, la del grupo de sociólogos del conocimiento que trabajan en el “programa fuerte”, cuyo punto de vista coincide con la opinión que no pocos científicos tienen sobre el papel de la filosofía de la ciencia, opinión según la cual los análisis filosóficos no tendrían nada que aportar en un programa interdisciplinario de estudios sobre la ciencia. Los defensores de dicho programa sostienen, por su parte, que la sociología es el mejor camino para alcanzar una comprensión científica de la ciencia misma. El problema básico, según estos autores, es el problema de explicar las creencias en términos de factores o mecanismos causales, explicación que no necesita tomar en cuenta las propiedades epistémicas de las creencias. Es decir, en la explicación de por qué un grupo de sujetos acepta o rechaza ciertas creencias no son relevantes las consideraciones sobre la justificación, la verdad o la objetividad de las mismas. Y se supone que el mismo tipo de mecanismos causales, como por ejemplo los ejercicios de poder entre los sujetos o grupos involucrados, ha de explicar todo tipo de creencias vigentes en una comunidad, al margen de los criterios epistemológicos para calificar una creencia como conocimiento. Estos sociólogos piensan que hay algo básicamente equivocado en los modelos epistemológicos, según los cuales existen contextos donde las creencias se aceptan con base en razones que son independientes de los intereses personales, posición social e ideología de los sujetos involucrados. Esta pretensión de racionalidad se considera un mito inventado por los filósofos, ya que, según su juicio, todos los casos de aceptación y cambio de creencias, considerados racionales o no, deben ser explicados por su vinculación causal con los factores sociales. De aquí el rechazo de los análisis epistemológicos, y en particular de los análisis filosóficos del conocimiento científico.
Son varias las críticas que un filósofo puede hacer a este tipo de programas de investigación sobre la ciencia, y por razones que van más allá de la mera lucha territorial. Una de las críticas más obvias al “programa fuerte” va dirigida contra su tendencia reduccionista. Los sociólogos de este programa cometen una falacia, pues reconocer que la ciencia es un fenómeno social no implica que la mejor —y menos la única— manera de dar cuenta de la ciencia sea en términos de factores y condicionamientos sociales. El innegable carácter social de esta empresa no justifica semejante pretensión, como tampoco lo haría la innegable dimensión social de una enfermedad como el sida, cuya explicación no se agota en los factores sociales involucrados en su propagación. Esta línea de crítica se aplicaría a cualquier teoría sobre la ciencia que pretendiera tener la perspectiva fundamental o privilegiada. Por otra parte, los sociólogos radicales del conocimiento, al estipular los requisitos que debe cumplir una explicación de las creencias para calificar ella misma como explicación científica, de hecho están adoptando ciertos criterios epistémicos sobre lo que cuenta como una buena explicación y sobre lo que constituye un enfoque científico. Por tanto, resulta incoherente que nieguen la importancia de este tipo de criterios filosóficos, cuando justo son aquello que justificaría sus pretensiones de conocimiento acerca del conocimiento. Todo estudioso de la ciencia —sea del campo que sea— que utilice criterios normativos en su quehacer, pero que a la vez niegue la utilidad o pertinencia de los análisis epistemológicos, caería en una inconsistencia semejante. Esta forma de argumentar en favor de la epistemología, cuyo problema central se podría formular como el problema del control de calidad de nuestras creencias y prácticas cognitivas, supone que existen problemas propiamente filosóficos que no se pueden reducir o asimilar a los planteados en la agenda de otras disciplinas. En el caso del conocimiento existe un área de la filosofía profesional que se ocupa expresamente de analizar los criterios que permiten distinguir aquellas afirmaciones o prácticas que son aceptables, confiables o razonables, de las que no lo son. Y cabe notar que tanto en la vida cotidiana como en la investigación científica, lo tengamos o no presente, hacemos constantemente ese tipo de distinciones y evaluaciones epistémicas. Un crítico de la epistemología podría replicar que él no tiene ningún reparo en aceptar que, en efecto, existen problemas de carácter filosófico en relación con las creencias y prácticas científicas, pero que esos problemas sólo pueden ser abordados seriamente por los mismos científicos. En pocas palabras, la filosofía de la ciencia sólo pueden hacerla los propios científicos. La respuesta a este crítico —que entre otras cosas estaría ignorando la utilidad de la división del trabajo— sería que ciertamente la reflexión filosófica exige un conocimiento a fondo de su objeto de estudio. Ningún filósofo de la ciencia pondría esto en duda. Pero tener un buen conocimiento de un campo de investigación no exige ser un científico practicante. La idea de este crítico, tomada en serio, equivaldría a afirmar que sólo los huicholes son capaces de estudiar sus ritos, o que sólo quienes van a misa y comulgan son capaces de estudiar el dogma católico. Lo cual, en algunos casos, ni siquiera resulta conveniente. Además, el hecho de que alguien sea un científico practicante tampoco garantiza que esté en mejor posición para tratar los problemas filosóficos de su disciplina. El análisis filosófico, como el trabajo científico, requiere de un entrenamiento profesional en el que se adquieren ciertas habilidades, destrezas y herramientas específicas. Sin embargo, por otra parte, también es preciso reconocer que algunas de las objeciones que se han hecho a la epistemología y la filosofía de la ciencia tradicionales tienen un trasfondo de razón. Al respecto, una de las repercusiones de mayor alcance del trabajo de Thomas Kuhn ha sido su contribución a una nueva manera de entender la filosofía de la ciencia, una manera que se ha catalogado como “naturalizada”. Si bien esta orientación tiene antecedentes importantes dentro del mismo campo de la filosofía, uno de los principales motores de este cambio de rumbo se encuentra en la obra más discutida de Kuhn, La estructura de las revoluciones científicas, publicada en 1962. A partir de que la filosofía de la ciencia se constituye como una disciplina académica especializada —alrededor de los años veintes de nuestro siglo— dominó el supuesto de que era posible descubrir y codificar los principios epistemológicos que gobernaban la actividad científica. Pero además se consideraba que dichos principios debían ser autónomos o independientes de la ciencia misma, pues de lo contrario no podrían fungir como fundamento de la evaluación y elección de teorías, ni de las normas del proceder científicamente correcto. Tales principios supuestamente constituían el núcleo de una racionalidad también autónoma -incondicionada y categórica- que estaba por encima de las prácticas y resultados de esta empresa cognitiva. Aunque Kuhn nunca utiliza el término “naturalización” para caracterizar la orientación de sus análisis —término que se vuelve de uso común a partir del trabajo de Quine publicado en 1969, de hecho éstos encierran el núcleo de lo que hoy se entiende por “epistemología naturalizada”. En contraste con el enfoque tradicional, se parte de la idea de que no hay un conjunto de normas o principios autónomos, pues ahora se considera que la epistemología no es independiente de la ciencia. Pero esto —por lo menos en el caso de Kuhn— no significa negar que hay mejores y peores maneras de hacer ciencia, ni rechazar la posibilidad de que el análisis epistemológico permita formular recomendaciones de procedimiento o juicios de valor sobre esta actividad (por ejemplo, sobre el carácter racional de casos concretos de aceptación o rechazo de teorías). Pero sí implica que este tipo de normativa y evaluación crítica se debe contextualizar tomando en cuenta la manera en que los agentes llevan a cabo su quehacer, es decir, lo que para ellos significa “hacer ciencia”, lo cual ciertamente ha variado en las distintas comunidades y periodos históricos. La epistemología tradicional —cuyo principal cuestionamiento ha provenido del análisis de la ciencia— requiere principios autónomos debido a su compromiso con una concepción demasiado estricta de la justificación de creencias, donde ésta debe proceder de manera lineal y en una sola dirección. Esto es, se considera que la justificación debe partir de principios “autoevidentes” o “autojustificatorios”, pues de lo contrario se correría el peligro de caer en un regreso al infinito o en una circularidad viciosa. De aquí que la justificación de las afirmaciones que se hacen en la ciencia deba apelar, en última instancia, a principios que sean por completo independientes de cualquiera de esas afirmaciones. En particular, lo que las teorías científicas digan sobre los procesos de percepción, aprendizaje o procesamiento de información, sobre la evolución de las diversas creencias y prácticas, sobre los grupos donde se generan y avalan los productos de investigación, etcétera, no puede ser tomado en cuenta en la justificación de dichas teorías. En una palabra, aquello que requiere justificación nunca podrá utilizarse para justificar o evaluar. En contraste con esta concepción tradicional, resalta un sentido muy básico en que el modelo de Kuhn del desarrollo científico implica una naturalización de la epistemología: los estándares de evaluación que operan en la ciencia no son del todo autónomos respecto de las teorías sobre el mundo, lo cual se puede generalizar afirmando que los estándares o criterios utilizados en las distintas comunidades científicas se modifican en función de la misma dinámica de la investigación. Kuhn destaca que los cambios de teoría, los cambios en el nivel de las afirmaciones empíricas, han repercutido en el nivel de los criterios de evaluación o justificación. Aquí vale la pena citarlo extensamente: “Lo que puede parecer especialmente problemático acerca de cambios como éstos [cambios en los criterios epistémicos] es, desde luego, que por lo regular ocurren como secuela de un cambio de teoría. Una de las objeciones a la nueva química de Lavoisier era que obstaculizaba el logro de aquello que hasta entonces había sido uno de los objetivos de la química tra dicional: la explicación de cualidades, como el color y la textura, así como el cambio de éstas. Con la aceptación de la teoría de Lavoisier tales explicaciones dejaron de ser, por algún tiempo, un valor para los químicos; la habilidad para explicar los cambios de cualidades dejó de ser un criterio relevante para evaluar una teoría química”. Sin embargo, “la existencia de un circuito de retroalimentación a través del cual el cambio de teorías afecta a los valores que condujeron a tal cambio, no hace que el proceso de decisión sea circular en ningún sentido nocivo”.
Este sentido en que Kuhn naturaliza la epistemología, al afirmar que los valores o estándares epistémicos son afectados y modificados por la dinámica de las teorías empíricas, va acompañado de una naturalización del análisis filosófico de la ciencia. Este análisis requiere ahora de la información que generan otros estudios sobre la ciencia, estudios que son de carácter empírico. La naturaleza social, histórica y evolutiva de la empresa científica, la cual se reconoce ampliamente a partir de Kuhn, así como la importancia que éste otorga a los aspectos psicológicos de ciertos procesos cognitivos, descubren una red de relaciones entre las diversas disciplinas donde se toma a la ciencia como objeto de estudio: la filosofía, la historia, la sociología, la psicología cognitiva y, más recientemente, la biología evolucionista. La tarea de establecer la naturaleza de estas relaciones, central para un programa interdisciplinario de estudios sobre la ciencia, apenas está en marcha —se podría decir que arranca en los años setentas—, y como se dijo proliferan las discusiones sobre la primacía de alguna de estas disciplinas frente a las demás. Si bien aquí no es posible entrar en esta intrincada discusión acerca de las relaciones de complementación, presuposición o reducción entre los diversos estudios metacientíficos, vale la pena mencionar las principales posiciones en el campo de la naturalización de la epistemología. La más radical es la posición que afirma que la epistemología debería ser reemplazada o sustituida por una ciencia empírica de los procesos cognitivos, y según se conciban estos procesos se propone a la psicología (como en el caso de Quine), a la sociología (como hacen los sociólogos del programa fuerte encabezados por Bloor), o a la biología evolucionista (como, por ejemplo, propone Campbell).
También encontramos posiciones integradoras que intentan combinar los resultados de ciertas ciencias empíricas con el análisis conceptual o filosófico, considerando que la investigación empírica sobre los sujetos y procesos epistémicos es una condición necesaria para comprender la cognición humana, pero que, recíprocamente, las ciencias empíricas requieren de un análisis y justificación de sus propios presupuestos, esto es, requieren de la epistemología. Esta sería la posición de un autor como Shimony. En esta misma línea encontramos modelos de interacción más dinámicos, donde se argumenta que los cambios en las formas o estrategias de investigación conducen a cambios en los estándares de evaluación, y que a su vez las nuevas consideraciones epistemológicas inciden en los programas de investigación científica planteando nuevos retos o generando nuevas preguntas. El modelo de Kitcher es un buen ejemplo de este tipo de propuesta.
Frente a las naturalizaciones radicales que proponen eliminar la epistemología, conviene insistir en la objeción de que sus programas de investigación sobre el conocimiento no se libran de los problemas epistemológicos tradicionales. Todos ellos parten del compromiso con determinados modelos de explicación, con cierto tipo de entidades teóricas, con alguna noción de verdad, con cierta concepción de la inferencia, etcétera, que son compromisos que requieren ser justificados. Como observa Shimony, si este tipo de teóricos del conocimiento se abstuviera de asumir supuestos normativos, su situación simplemente sería análoga a la de alguien que “deposita toda su confianza en los matemáticos y asume la corrección de sus útiles teoremas, sin revisar las pruebas él mismo”. Pero dado que la mayoría de ellos intentan lidiar con cuestiones normativas, no pueden evitar el problema de la justificación de sus propios postulados. Por otra parte, el reto para los enfoques naturalizados que intentan preservar una función normativa o evaluativa para la epistemología, es en qué sentido y en qué medida pueden dar cuenta de la racionalidad del desarrollo del conocimiento.
Al respecto, cabe decir que la reticencia que se observa en las últimas publicaciones de Kuhn hacia los enfoques naturalizados se explicaría por su rechazo de las versiones extremas o reduccionistas, las cuales chocaban con su inmersión cada vez mayor en el análisis filosófico de la ciencia. El interés creciente de Kuhn en cuestiones como la racionalidad, el relativismo, la verdad y el realismo —temas centrales del libro que no llegó a publicar en vida— tenía que contraponerlo a las propuestas que eliminan como irrelevante a la filosofía de la ciencia.
Sin embargo, está claro que para este autor la racionalidad que opera en la actividad científica no es autónoma ni categórica. Los criterios de evaluación de hipótesis y teorías —criterios como simplicidad, precisión, consistencia, alcance, etcétera—, además de sufrir transformaciones en su interpretación y jerarquía a través del desarrollo de las distintas disciplinas, tienen siempre un carácter condicional o instrumental, esto es, conectan las estrategias de investigación con los objetivos perseguidos. Un científico actúa de manera racional cuando elige y utiliza los medios más efectivos, de entre los disponibles, para alcanzar las metas deseadas. También queda claro que, de acuerdo con el modelo de Kuhn, la tarea filosófica de evaluar la racionalidad de un cambio de teoría requiere la información que proporciona la investigación empírica sobre la ciencia. Es necesario apoyarse en estudios detallados del contexto de investigación y descubrir, en cada periodo del desarrollo de una disciplina, cuáles eran los objetivos, los supuestos, los procedimientos experimentales, las herramientas formales y los criterios de evaluación vigentes, para explicar que los científicos en cuestión hayan considerado una teoría como mejor que otra, y poder juzgar, en su contexto, si ese cambio de enfoque teórico fue un cambio racional o razonable. Esta vinculación entre filosofía e investigación empírica no sólo pone en tela de juicio la idea de que la filosofía de la ciencia se basa en principios autónomos y tiene un carácter puramente normativo. También pone en cuestión la idea de que el epistemólogo que adopta una perspectiva naturalizada se limita a describir lo que los científicos de hecho creen o hacen. Como señala Ronald Giere, una filosofía naturalizada de la ciencia —bien entendida— es semejante a una teoría científica en el sentido de que ofrece algo más que meras descripciones. En ambos casos hay una base teórica que no sólo permite elaborar explicaciones sobre su objeto de estudio, sino que también permite orientar la forma en que se conduce la investigación. Esto es, las teorías, en general, proporcionan una base para formular juicios normativos y evaluativos. Cuando Feyerabend cree detectar en el trabajo de Kuhn una ambigüedad a este respecto, dice: “Siempre que leo a Kuhn, me surge la siguiente pregunta: ¿estamos ante prescripciones metodológicas que dicen al científico cómo proceder, o frente a una descripción, vacía de todo elemento evaluativo, de aquellas actividades que generalmente se llaman ‘científicas’”? La respuesta de Kuhn va justamente en la línea de naturalización recién apuntada: “Si tengo una teoría de cómo y por qué funciona la ciencia, dicha teoría necesariamente tiene implicaciones sobre la forma en que los científicos deberían comportarse si su empresa ha de prosperar”. Esta respuesta no sólo rompe con la dicotomía entre lo prescriptivo y lo descriptivo sino que también revela que los juicios normativos sobre la actividad científica, además de depender de una teoría sobre la ciencia, tienen siempre un carácter condicional o instrumental. El esquema de la posición kuhniana sería como sigue: los científicos se comportan de tales y cuales formas; algunas de esas formas cumplen ciertas funciones básicas (es decir, permiten el logro de ciertos objetivos); en ausencia de formas alternativas que cumplieran funciones similares, los científicos deberían comportarse de tales maneras si su objetivo es hacer avanzar el conocimiento científico. Desde un enfoque naturalizado, ésta es la única clase de prescripciones legítimamente formulables, dado que no hay una racionalidad absoluta (ni autónoma ni categórica) como suponía la mayoría de filósofos de la ciencia anteriores a Kuhn. |
|||
|
Referencias Bibliográficas
Bloor, D. 1976, Knwoledge and Social Imagery, Londres, Routledge Kegan Paul. Feyerabend, P. 1970, “Consuelos para el especialista”, en I. Lakatos y A. Musgrave (eds.), La crítica y el desarrollo del conocimiento, Barcelona, Grijalbo, 1975, pp. 345-389. Giere, R. l989, “Scientific Rationality as Instrumental Rationality”: Studies in History and Philosophy of Science, Vol. 20, No. 3, pp. 377-384. Kitcher, P. 1993, The Advancement of Science. Science without Legend, Objectivity without Illusions, New York, Oxford University Press. Kuhn, T.S. 1962, La estructura de las revoluciones científicas, México, Fondo de Cultura Económica, 1971. Kuhn, T.S. 1970, “Consideración en torno a mis críticos”, en I. Lakatos y A. Musgrave (eds.), La crítica y el desarrollo del conocimiento, Barcelona, Grijalbo, 1975, pp. 391-454. Kuhn, T.S. 1977, “Objetividad, juicios de valor y elección de teorías”, en La tensión esencial, México, CONACYT/FCE, 1982, pp. 320-339. Quine, W.V.O. 1969, “Epistemology Naturalized”, en Ontological Relativity and Other Essays, New York, Columbia University Press, pp. 69-90. Shimony, A. 1993, Search for a Naturalistic World View, Vol. I, Cambridge, Cambridge University Press. |
|||
|
Ana Rosa Pérez Ransanz
Instituto de Investigaciones Filosóficas,
Universidad Nacional Autónoma de México.
_______________________________________________________________ como citar este artículo → Pérez Ransanz, Ana Rosa. (1999). Thomas S. Kuhn y la "naturalización" de la filosofía de la ciencia. Ciencias 53, enero-marzo, 44-49. [En línea]
|
|||
| ← Regresar al índice ⁄ artículo siguiente → |