
")

 |
 |
|
| La construcción de la doble hélice, de la nucleína al adn |
|
|
|
Martín Bonfil Olivera
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Es la molécula la que tiene glamour,
no los científicos.
Francis Crick
|
||
|
Pocas veces un hallazgo científico marca de manera tajante el inicio de una nueva era. Es el caso del famoso artículo firmado por J. D. Watson y F. H. C. Crick en el número del 25 de abril de 1953 de la revista inglesa Nature. Con sólo 14 párrafos, el escueto título de “Molecular structure of nucleic acids. A structure for desoxyribose nucleic acid” y una sola figura —que luego llegaría a convertirse en un icono: la “escalera de caracol” de la doble hélice del adn—, el artículo de Watson y Crick bastó para marcar formalmente el nacimiento de la genética molecular. Constituye, junto con la formulación de la teoría de la selección natural de Darwin, o de la teoría celular por Schleiden y Schwann, uno de los hitos decisivos en la historia intelectual de las ciencias biológicas.
Al igual que la memoria, la historia de la ciencia tiende a simplificar los hechos, a veces hasta el punto de caricaturizarlos. Los descubrimientos científicos, vistos a la distancia, parecen logros de genios que conciben de pronto grandes ideas que resultan, casi por casualidad, correctas. Pero rara vez, al menos en las ciencias naturales, se puede hablar de descubrimientos individuales. La metáfora de los hombros de gigantes puede estar ya desgastada, pero no por ello es menos cierta. Al igual que las especies biológicas, las ideas no surgen espontáneamente de la nada: evolucionan a partir de ideas anteriores, generadas en otros cerebros. También mutan y se combinan unas con otras; producen descendencia variada que luego es seleccionada, en parte, por su capacidad para colonizar nuevas mentes, pero sobre todo, en el caso de la ciencia, para ajustarse a eso que llamamos realidad.
El descubrimiento de la doble hélice surge, principalmente, de dos linajes de ideas. Uno proviene de la genética y se remonta a los conceptos de los antiguos griegos sobre la pangénesis, que suponía que ciertas secreciones del cuerpo de los padres se mezclaban para dar origen a un hijo. El otro son los estudios acerca de la química del material hereditario.
En el siglo XIX hubo varias teorías que buscaban explicar la transmisión de caracteres hereditarios de padres a hijos. Una de ellas, un refinamiento de la pangénesis, fue desarrollada por Charles Darwin. Pero la genética sólo se formalizó como estudio científico con los trabajos del monje austriaco Gregor Mendel, publicados en 1866 en la oscura revista de la Sociedad de Ciencias Naturales de Brno. En ellos demostraba la existencia de unidades hereditarias que se transmitían de padres a hijos sin mezclarse y siguiendo unas leyes sencillas. Desgraciadamente los estudios de Mendel no fueron tomados en cuenta por los científicos de su tiempo. Permanecieron olvidados durante cuarenta y cuatro años, hasta que en 1900 fueron redescubiertos simultánea e independientemente por tres botánicos que habían reproducido sus experimentos: el holandés Hugo de Vries, el alemán Karl Correns y el austriaco Erich von Tschermak.
A partir de ese momento fue quedando cada vez más claro que para entender a fondo la herencia había que desentrañar el funcionamiento de estas unidades que el botánico danés Wilhelm Johannsen denominaría genes en 1909.
De Vries había estado trabajando algún tiempo sobre la herencia, antes de redescubrir los hallazgos de Mendel, y en 1889 formuló una teoría llamada pangénesis intracelular, notoria porque prefigura varias de las características de la moderna genética molecular. Hacia 1850 no estaba todavía claro que el núcleo de la célula fuera importante para la vida, pero en 1866 el prestigiado naturalista alemán Ernst Haeckel reconoció que “el núcleo interno permite la transmisión de caracteres hereditarios”, lo cual contribuyó a afianzar la noción de que los factores responsables de la herencia debían estar localizados en el núcleo (posteriormente, alrededor de 1910, los trabajos del zoólogo estadounidense Thomas Hunt Morgan comprobarían que se encuentran más precisamente en los cromosomas).
De Vries destacó en su teoría esta localización nuclear del material hereditario. “Considero que el resultado más importante de la investigación celular de la década pasada —escribe en 1889— es la teoría de que todas las predisposiciones hereditarias del organismo deben estar representadas en el núcleo de la célula”. Asimismo, en un párrafo que parece predecir los descubrimientos moleculares de décadas futuras, postula que “los portadores materiales de los caracteres hereditarios no pueden ser idénticos a las moléculas químicas; deben concebirse como unidades, construidas a partir de éstas, mucho más grandes que ellas y aún así invisiblemente pequeñas”.
Sin embargo, al llegar a este punto, poco podía hacerse para averiguar el funcionamiento de los genes. Era necesaria otra línea de ataque para descifrar el enigma. A pesar de que los estudios de genética clásica seguirían proporcionando importantes descubrimientos, como el mapeo cada vez más preciso de genes en los cromosomas y la identificación de genes relacionados con funciones o enfermedades específicas, fue un segundo linaje conceptual el que llevó finalmente a conocer la estructura del adn. Se trata del que proviene de los estudios de la química de los sistemas vivos, iniciado con la síntesis de las primeras moléculas orgánicas en laboratorio (la obtención de la urea por Wöhler en 1828), continuada por el nacimiento y desarrollo de la bioquímica hasta llegar a la biología molecular propiamente dicha. Los detalles rara vez se mencionan al hablar de la historia del adn.
El padre del adn
El ácido desoxirribonucleico fue descubierto por Friedrich Miescher, bioquímico suizo nacido en 1844 y muerto en 1895. En 1869 Miescher trabajaba en Tubinga haciendo “histoquímica”. En una época en la que muchos investigadores se dedicaban a examinar el funcionamiento de la célula, Miescher decidió concentrarse en el estudio de células linfoides (glóbulos blancos o linfocitos), mismas que podían obtenerse fácilmente a partir de la pus de los enfermos de una clínica cercana, pues entonces las infecciones graves eran cosa común.
Miescher estaba interesado en la composición química de las células, en particular en las proteínas, sustancias presentes en el “protoplasma” (la “sustancia viva” en el interior de la célula, equivalente a lo que hoy conocemos como citoplasma, junto con los organelos que contiene). Por ello se dedicó a tratar de separar el protoplasma y los núcleos de las células. Buscando cómo lograrlo aplicó diferentes soluciones de sales a las células mientras las observaba en el microscopio. Fue entonces que hizo una observación importante que desviaría su atención del protoplasma al núcleo: “En el experimento con fluidos débilmente alcalinos, se obtuvieron a partir de las soluciones, mediante neutralización, precipitados que no eran solubles en agua, ácido acético, ácido clorhídrico muy diluido ni en solución de cloruro de sodio, y en consecuencia no pueden pertenecer a ninguna de las sustancias proteicas conocidas hasta ahora”.
Miescher tenía razones para suponer que este precipitado provenía de los núcleos de las células, lo cual era la primera indicación de que el núcleo pudiera tener una composición química propia. Para comprobarlo tuvo que desarrollar una técnica para aislar los núcleos, lo cual logró utilizando pepsina (enzima que destruye proteínas, obtenida de extractos de estómago de cerdo) para romper las células. Una vez obtenidos los núcleos en forma pura (lo cual comprobó mediante el microscopio), aisló nuevamente el precipitado a partir de ellos.
Posteriormente Miescher también logró purificar su misteriosa sustancia a partir de núcleos de levaduras, células de riñón, de hígado, de testículo y glóbulos rojos nucleados y confirmó que no parecía tratarse de una proteína. Tomando en cuenta su origen decidió llamarla nucleína.
El análisis químico indicaba que, además de carbono, oxígeno, nitrógeno e hidrógeno, la nucleína contenía fósforo, un elemento poco usual en las sustancias de origen biológico estudiadas hasta entonces. El contenido de fósforo en la nucleína fue lo que contribuyó en gran medida a despertar el interés de otros investigadores en su estudio. Por diversas circunstancias, entre ellas una guerra, los hallazgos de Miescher no se publicaron sino hasta 1871.
Inicialmente Miescher pensaba que la función de la nucleína era servir como reserva de fósforo, manteniéndolo disponible para cuando la célula lo necesitara. Mediante el uso de esperma de salmón, una excelente fuente de núcleos, continuó estudiando la composición química de esta sustancia, aunque la labor se complicaba por lo difícil que resultaba separar la nucleína de las proteínas que, según descubrió el propio Miescher, también estaban presentes en el núcleo. Hacia el final de su vida Miescher sospechaba, junto con otros, que la nucleína podía constituir la base química de la herencia.
En 1889 Richard Altmann reportó la obtención de nucleína libre de proteínas, con lo cual comprobó que éstas últimas no contenían fósforo y sugirió, tomando en cuenta su carácter ácido, un nuevo nombre para la nucleína: ácido nucleico.
El núcleo y los cromosomas
Mientras tanto los estudios del núcleo celular habían seguido avanzando paralelamente. Entre 1870 y 1880 el bacteriólogo alemán Paul Ehrlich había desarrollado técnicas de tinción química que le permitieron distinguir diversos componentes celulares, entre ellos los cromosomas, llamados así por su propiedad de teñirse con colorantes básicos. En la misma década varios investigadores se dedicaron a describir la estructura y comportamiento de los cromosomas durante la división celular. Para 1881 Eduard Zacharias aplicó las técnicas desarrolladas por Miescher al estudio de los cromosomas, disolviendo con pepsina el citoplasma de las células y dejando núcleos aislados. Los cromosomas resistían la acción de la pepsina, lo cual indicaba que su naturaleza no era proteica.
En 1879 el alemán Walther Flemming propuso el término cromatina para denominar al material intensamente coloreado que aparecía en el núcleo en interfase (cuando no se está dividiendo, a diferencia de los cromosomas, que aparecen durante la división) al teñirlo con colorantes básicos. En un estudio de 1882 Flemming escribe: “En vista de su naturaleza refringente, sus reacciones, y sobre todo su afinidad por los colorantes, [he nombrado “cromatina” a esta sustancia]. Posiblemente la cromatina es idéntica a la nucleína, pero si no, puede deducirse de los estudios de Zacharias que la una contiene a la otra. La palabra cromatina puede servirnos hasta que su naturaleza química sea conocida, y mientras tanto significa la sustancia en el núcleo celular que se tiñe fácilmente”. Comenzaba así a converger esta línea de investigación citológica con la de Miescher, que era más bioquímica.
¿De qué está hecha la nucleína?
La composición química de la nucleína era un problema central en el camino que llevaría directamente al descubrimiento de la doble hélice, pero antes de abordar la estructura de una molécula tan complicada tuvo que estudiarse su composición en términos de las unidades más sencillas que la forman.
En 1879 el bioquímico alemán Albrecht Kossel realizó análisis químicos de la nucleína de levaduras (hongos unicelulares que en esa época eran considerados plantas) y encontró en su composición, además de fósforo en forma de fosfato, un compuesto heterocíclico del tipo de las bases nitrogenadas (con anillos de carbono y nitrógeno) llamado guanina. Posteriormente descubrió también la presencia de dos nuevas bases, mismas que llamó adenina y timina. La citosina sería descubierta poco después. En 1893 Kossel descubrió que los ácidos nucleicos contenían también un carbohidrato, que identificó como una pentosa (azúcar de cinco carbonos). En 1909 esta pentosa fue identificada con más precisión como la d-ribosa. Paralelamente, también realizó estudios que mostraron que el papel de la nucleína en células de hígado y bazo, más que de reserva, se relacionaba con la formación de nuevos tejidos.
Durante un tiempo se pensó que el ácido nucleico de timo de ternera contenía otro tipo de azúcar, una hexosa (seis carbonos), pero posteriormente, mediante estudios muy cuidadosos, el químico ruso Theodor Levene y sus colaboradores encontraron que en realidad también se trataba de una pentosa: la 2-desoxi-d-ribosa. Así, durante algún tiempo se creyó que los ácidos nucleicos de las plantas contenían ribosa y los de los animales desoxirribosa. Hoy sabemos que el ácido nucleico de ribosa o arn es especialmente abundante en las levaduras, mientras que el de desoxirribosa o adn abunda en el timo.
Establecidos así los componentes más sencillos de los ácidos nucleicos, comenzó a elucidarse el siguiente nivel de su estructura.
En 1909 Levene y su colaborador Jacobs determinaron que la base nitrogenada y el fosfato están unidos respectivamente a los carbonos 1 y 5 de la pentosa. Propusieron los términos nucleósido y nucleótido para denominar respectivamente a la unión de una purina y un carbohidrato y a su éster fosfórico.
Ya desde 1893 Kossel y otros habían estudiado con métodos bastante burdos las proporciones de las purinas y pirimidinas que se hallaban en los ácidos nucleicos. En el ácido nucleico de timo las cuatro parecían hallarse en cantidades iguales. Esto hizo que surgiera la que hasta la década de 1940 sería la hipótesis dominante: que la unidad fundamental de los ácidos nucleicos era un “tetranucleótido” que contenía una de cada una de las cuatro bases (incluso, en 1935, llegó a proponerse que se trataba de tetranucleótidos cíclicos).
Para 1938 había quedado claro que los dos tipos de ácido nucleico, arn y adn, se hallaban presentes tanto en plantas como animales, pero la investigación sobre su estructura se hallaba bastante estancada. Una dificultad importante era determinar el peso molecular de los ácidos nucleicos. En la década de los veintes no se sabía si se trataba de coloides, agregados de moléculas pequeñas unidas por enlaces débiles, o polímeros, moléculas gigantes unidas por enlaces covalentes. Hasta ese momento los métodos usados para aislarlo eran más bien burdos, lo que provocaba su ruptura. Se obtenían pesos moleculares de alrededor de 1 500, cercanos a lo esperado para un tetranucleótido, por lo cual esta hipótesis se vio reforzada.
En 1938, mediante cuidadosos estudios que recuperaban técnicas desarrolladas por Miescher, el sueco Einar Hammarsten preparó y estudió adn de timo (“ácido timonucléico”) y determinó mediante un método óptico que éste tenía un peso molecular muy alto (entre 500 mil y un millón), mismo que era de 300 a 600 veces mayor que el de las proteínas. Este hallazgo se confirmó mediante otros métodos como la ultracentrifugación y la difracción de rayos X. El alto peso molecular implicaba que se trataba de un polímero, quizá un polímero de tetranucleótidos. Sin embargo no parecía tener un peso molecular bien definido, por lo cual se trataba de una molécula extraña.
Volviendo a la química básica
La química es una ciencia que tiene dos caras. Una de ellas es el análisis, vía seguida hasta entonces en el estudio de los ácidos nucleicos. La otra es la síntesis, que permite confirmar la estructura que se ha deducido para una molécula. En 1947 el químico escocés Alexander Todd consiguió sintetizar dos nucleótidos, el mono y el difosfato de adenosina (amp y adp), logro que confirmó la estructura de estas moléculas y por el cual en 1957 recibiría el premio Nobel.
En el mismo año quedó claro que la diferencia entre el arn y el adn era la ausencia en este último del grupo hidroxilo en el carbono 2’ de la ribosa (2-desoxirribosa, de ahí su nombre actual de ácido desoxirribonucleico). Es precisamente la presencia de este hidroxilo en el arn lo que le confiere una mayor reactividad, y por tanto una menor estabilidad, lo cual probablemente explique porqué, a pesar de que se piensa que las primeras moléculas autorreplicantes eran arn, los organismos actuales tienen un genoma de adn y no de arn.
Entre tanto, la hipótesis de los tetranucleótidos comenzó a perder fuerza. Esto se debió en parte a los trabajos del químico inglés John Masson Gulland, quien en 1947, mediante titulaciones ácido-base muy precisas, estudió la estequiometría del adn y mostró que la cantidad de grupos fosfato ionizables por cada molécula era superior a la que correspondería a un tetranucleótido. Adelantándose al futuro modelo de Watson y Crick, Gulland también mostró, mediante el mismo método, que parecía haber grupos débilmente ionizables en las bases que podrían estar formando enlaces de hidrógeno entre ellas (aunque no se podía saber si unían partes de una misma cadena o cadenas distintas de adn).
El último gran aporte de la química tradicional en la ruta hacia la doble hélice fue el trabajo realizado por el bioquímico austriaco Erwin Chargaff, quien entre 1948 y 1952 estudió, mediante cromatografía en papel, las proporciones de las cuatro bases en el adn. Chargaff halló que los porcentajes de bases presentes en diversas especies eran muy variables, aunque la cantidad de purinas totales, adenina y guanina, era siempre igual a la cantidad de pirimidinas, timina y citosina. Éste fue el tiro de gracia para la hipótesis de que el adn estaba formado por tetranucleótidos que contenían una base de cada clase. Sin embargo su mayor hallazgo fueron las famosas reglas de Chargaff, producto de una medida refinada de la estequiometría de las bases en el adn de especies distintas: la cantidad de adenina es siempre equivalente a la de timina y la de guanina a la de citosina. Ésta fue, como se sabe, una de las pistas clave que permitieron a Crick y Watson concretar su modelo.
El resultado de todo este trabajo químico es resumido así por Portugal y Cohen: “a principios de los cincuenta se sabía que el adn era un polímero de alto peso molecular en el que grupos fosfato unían a los desoxirribonucleósidos entre las posiciones 3’ y 5’ de sus azúcares. La secuencia de las bases se desconocía, aunque se habían observado algunas regularidades cuantitativas en la composición de bases. Aun cuando la estructura química detallada del adn había sido determinada, su geometría molecular permanecía en el misterio”. El próximo paso sería la transición a la biología molecular, historia que se narra con más frecuencia.
Nace la genética molecular
Paralelamente a la investigación química del adn, los estudios genéticos estaban demostrando el papel central de esta molécula como agente hereditario.
En 1928 el bacteriólogo inglés Frederick Griffith descubrió el fenómeno de transformación genética en las bacterias causantes de la neumonía (Pneumococcus). Se conocían dos formas del microorganismo: una patógena o forma S, capsulaza, y otra inocua o forma R, sin cápsula. Griffith halló que al inocular en ratones bacterias vivas de la forma R, junto con bacterias tipo S muertas por calentamiento, se observaba la transformación de las bacterias R en S vivas, mismas que eran capaces de causar la enfermedad.
En 1944, utilizando cuidadosas técnicas de purificación, el médico canadiense Oswald Avery, junto con Colin MacLeod y Maclyn McCarty, purificó el “principio transformante” responsable de este fenómeno y comprobó que se trataba de adn (por ejemplo, al mostrar que la enzima desoxirribonucleasa, que destruye al adn, eliminaba la actividad transformante). La cantidad de técnicas utilizadas para fundamentar su hallazgo se revela en un párrafo al final de su histórico artículo: “Los datos obtenidos mediante análisis químicos, enzimáticos y serológicos [inmunológicos], junto con los resultados de los estudios realizados por electroforesis, ultracentrifugación y espectroscopía ultravioleta indican que […] la fracción activa no contiene en forma demostrable proteína, lípido libre o polisacárido serológicamente reactivo, y consiste principal, si no exclusivamente, en una forma altamente polimerizada y viscosa de ácido desoxirribonucleico”.
Estos resultados, que enfocaron decisivamente la atención en el adn, no fueron aceptados como definitivos pues permanecía la posibilidad de que hubiera pequeñísimas cantidades de proteína que pudieran transmitir la información genética, como suponían todavía muchos investigadores.
Las dudas no se esfumarían hasta la publicación en 1952 de un trabajo firmado por Alfred Hershey y Martha Chase, quienes provenían de otra tradición experimental: la que se dedicaba al estudio de los virus de bacterias o bacteriófagos (fagos). Este tipo de estudios había logrado un alto grado de refinamiento, lo que permitió que Hershey y Chase hicieran el experimento decisivo para confirmar el papel genético del adn. Utilizando cultivos de fagos marcados radiactivamente, unos con azufre (que sólo está presente en las proteínas) y otros con fósforo (que se halla sólo en el adn del virus), comprobaron que cuando un fago infecta a una bacteria para reproducirse, sólo su adn penetra en ella y es suficiente para la multiplicación del fago. Con esto el papel genético del adn era ya indudable.
1951-1953: La odisea de Cambridge
Uno de los libros más citados al hablar de la historia del adn es ¿Qué es la vida?, escrito en 1944 por el físico Erwin Schrödinger. En él reflexiona sobre la constitución física del material genético: “A menudo se ha preguntado cómo, en esta diminuta mancha de materia, el núcleo de un óvulo fertilizado, puede estar contenida una clave elaborada y que contiene todo el desarrollo futuro del organismo. Una asociación bien ordenada de átomos, capaz de mantener permanentemente su orden, parece ser la única estructura material concebible que ofrece una variedad de posibles organizaciones (‘isoméricas’) y que es suficientemente grande como para poder contener un sistema complicado de ‘determinaciones’ dentro de reducidos límites espaciales”.
Schrödinger postulaba que el gen debía ser un “sólido aperiódico” y profetizaba (en forma finalmente errónea, pero sugestiva) que, “si bien [la materia viva] no elude las ‘leyes de la física’ tal como están establecidas hasta la fecha, probablemente implica ‘otras leyes físicas’ desconocidas hasta ahora”.
Sus ideas tuvieron el efecto de despertar en muchos el interés por el estudio molecular de los genes, en especial en el físico inglés Francis Harry Compton Crick y el biólogo estadounidense James Dewey Watson. “Un factor importante que contribuyó a su abandono [de Crick] de la física y al desarrollo de un acusado interés por la biología —comenta Watson en su relato autobiográfico La doble hélice— había sido la lectura, en 1946, de la obra del célebre físico teórico”.
Este libro también ejerció una influencia importante en el mismo Watson, quien lo descubrió en una biblioteca en el mismo año y dice en A passion for dna: “Después de leerlo, nunca fue lo mismo […] Que el gen fuera la esencia de la vida era claramente más importante que cómo migran las aves, el tema científico del que anteriormente nunca me saciaba […] Para finales del periodo lectivo, había decidido tener al gen como el principal objetivo de mi vida”.
El desarrollo de una nueva técnica física, la cristalografía por difracción de rayos X, permitió abordar el estudio de macromoléculas como las proteínas y el adn. En 1951, cuando Watson llegó al laboratorio Cavendish en Cambridge y conoció a Crick, ya había un grupo en el King’s College de Londres, encabezado por el físico neozelandés Maurice Wilkins, dedicado a aplicar esta técnica al análisis de la estructura molecular del adn. Crick y Watson, quien originalmente había tratado de colaborar con Wilkins, abordaron el estudio del adn informalmente. Quizá fue esto lo que les permitió tener un enfoque fresco que a la postre resultaría tan productivo.
El relato detallado de cómo la cristalografía de rayos X —junto con el caudal de conocimiento acumulado acerca de la química del adn y la técnica de construcción de modelos moleculares del químico estadounidense Linus Pauling— se aplicó para develar la estructura de la doble hélice, ya ha sido abundantemente narrado. El 28 de febrero de 1953, meses después de haber propuesto un primer modelo (erróneo) de tres cadenas, Crick y Watson construyeron una hélice formada por dos cadenas de adn corriendo en forma antiparalela. Las bases nitrogenadas formaban pares específicos en el centro, unidos por puentes de hidrógeno: la adenina se une a la timina y la guanina a la citosina. Se explicaban así las misteriosas reglas de Chargaff y al mismo tiempo se veía claramente cómo cada cadena podía servir de molde para construir otra idéntica, proporcionando así una base para entender la replicación del adn.
El logro de Watson y Crick, apoyado en los datos cristalográficos obtenidos por Wilkins y su colaboradora Rosalind Franklin, constituye el punto en que por fin se anudan los cabos de esta historia. Genética y bioquímica se trenzan con la biología celular para dar origen a la biología molecular, proporcionando así los fundamentos sobre los que Watson y Crick pudieron construir su famoso modelo. El proceso por el que evolucionan las ideas requiere la mezcla de muchos linajes; en este caso fue necesaria la aportación de numerosos cerebros durante más de ochenta años. No obstante, todavía faltaba tiempo para que el modelo fuera admitido por completo.
La aceptación de una idea
En su relato Watson hace énfasis en la competencia con el gran químico estadounidense Linus Pauling por descifrar la estructura del adn. Pauling había propuesto recientemente, con gran éxito, la estructura llamada hélice alfa, presente en proteínas fibrosas como la queratina, y había anunciado que abordaría el problema de la estructura del adn.
Pauling y su colaborador Robert B. Corey propusieron un modelo de triple hélice con los fosfatos al centro para el adn, muy similar a la fallida propuesta que habían hecho anteriormente Crick y Watson. En una carta dirigida a su hijo Peter, fechada el 18 de febrero de 1953 (exactamente diez días antes de que Crick y Watson dieran con la estructura acertada), Pauling escribe: “Me agrada tener noticias tuyas acerca de los ácidos nucleicos […] Oí un rumor de que Jim Watson y Crick habían formulado ya esta estructura hace algún tiempo, pero no habían hecho nada con ella. Probablemente el rumor era exagerado”.
El 27 de marzo de 1953, luego de recibir una copia del manuscrito del artículo que Watson y Crick habían enviado para su publicación, Pauling les contestó en los siguientes términos: “Estimados Dr. Watson y Mr. Crick: Estoy encantado de recibir su carta del 21 de marzo, y de ver la carta que están enviando a Nature. Creo que está bien que haya ahora dos estructuras propuestas para el ácido nucleico, y espero con ansia saber cuál será la decisión acerca de cuál es la incorrecta. Sin duda los datos del King’s College eliminarán una o la otra”.
Sin embargo, para el 20 de abril, cinco días antes de la publicación del artículo, la decisión ya estaba clara incluso para el propio Pauling, como muestra en una carta dirigida a Max Delbrück: “Quedé muy impresionado con la estructura de Watson-Crick. No sé si usted sabe lo que nos desvió a Corey y a mí al camino falso. Las fotografías de rayos X que teníamos […] son realmente la superposición de dos patrones, debido a dos diferentes modificaciones de los timonucleatos de sodio. Esto había sido descubierto hace un año o más por la gente del King’s College, pero no lo habían anunciado […] Aunque hay todavía una pequeña posibilidad de que su estructura [de Watson y Crick] esté equivocada, creo que es muy probable que sea correcta […] Creo que es el más importante avance que se ha dado en largo tiempo”.
Efectivamente, el modelo de la doble hélice tardó todavía varios años en quedar comprobado plenamente. Se manejaron diversas objeciones acerca de cómo el entrelazamiento de las cadenas dificultaría la replicación y se exploró la posibilidad de que en realidad se tratara de dos cadenas que corrían lado a lado sin estar entrelazadas (“yo no creí una palabra de esto —comenta Crick. Además, los modelos eran feos”). Todavía en 1979, Crick escribió con otros un artículo de revisión titulado “¿Es el adn realmente una doble hélice?” Como explica el propio Crick, “sólo hasta el comienzo de los ochentas quedó finalmente cofirmada la estructura en doble hélice del adn”.
La historia del adn es un excelente ejemplo de la complejidad del proceso de construcción del conocimiento científico. Los 50 años de su culminación nos hacen apreciar, nuevamente, la belleza no sólo de la estructura finalmente revelada, sino también la del proceso por el que paulatinamente se fue construyendo esta elegante imagen.
|
||
|
Referencias bibliográficas
Crick, Francis H. C. 1988. What mad pursuit. Basic Books, Nueva York.
Pauling, Linus. Papers at the U.S. National Library of Medicine (http://profiles.nlm.nih.gov/MM/Views/ Exhibit/documents/biomolecules.html)
Portugal, H. Franklin y Jack S. Cohen. 1977. A century of dna. mit Press, Cambridge, Mass.
Schrödinger, Erwin. 1944. ¿Qué es la vida? 2ª edición, Tusquets, Barcelona, 1983.
Watson, James D. 1968. La doble hélice. Plaza y Janés, Barcelona, 1970; también Conacyt, México, 1981.
Watson, James D. 2001. A passion for dna. Cold Spring Harbor Laboratory Press, Nueva York.
|
||
|
Martín Bonfil Olivera
Dirección General de Divulgación de la Ciencia, Universidad Nacional Autónoma de México.
_______________________________________________________________
como citar este artículo → Bonfil Olivera, Martín. (2003). La construcción de la doble hélice, de la nucleína al ADN. Ciencias 71, julio-septiembre, 4-15. [En línea] |
||
| ←Regresar al índice / artículo siguiente→ | ||
 |
 |
|
| Principales líneas de estudio en la biología evolutiva contemporánea |
|
|
|
Douglas J. Futuyma
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Actualmente vivimos un momento sumamente excitante en la biología evolutiva de los últimos cincuenta o sesenta años, y lo es simplemente porque ha habido un enorme progreso en casi cada aspecto del estudio de la evolución. Este progreso se debe a varios factores: por principio de cuentas se debe a avances tecnológicos tales como el poder secuenciar el adn, poder estudiar la expresión génica o la capacidad de alcanzar la transformación genética pasando genes de un organismo a otro. La aceleración en dicho progreso es el resultado de avances en la tecnología de la información. Hoy día contamos con bases de datos como los bancos de genes y poseemos una enorme capacidad de cómputo que hace posible resolver problemas como las relaciones filogenéticas de grandes grupos de organismos que no hubieran podido ser resueltas en el pasado. También existen avances en la teoría del cambio evolutivo, por ejemplo la teoría de la coalescencia, que es un aspecto enormemente importante y nuevo en genética de poblaciones. Incluso hay cambios en la forma en que podemos analizar los datos evolutivos. Hoy día también poseemos métodos estadísticos sofisticados para mapear los loci de rasgos cuantitativos (lrc o qtl por sus siglas en inglés), lo cual significa que podemos encontrar aquellos loci que contribuyen a la variación en las características poligénicas. Además existen cambios sociológicos que añaden avances a la biología evolutiva, por ejemplo hay una conjunción entre los estudios microevolutivos y los macroevolutivos.
Finalmente, también existen avances en la biología evolutiva simplemente porque se ha incrementado el poder de comunicación gracias al correo electrónico, a las revistas científicas en línea que hacen posible el intercambio entre biólogos de todo el mundo, y esto en sí mismo contribuye al avance de toda la ciencia. Y ya que el progreso ha tocado casi cada aspecto de la biología evolutiva, me concentraré en los aspectos de mayor interés personal.
Los estudios filogenéticos
La producción de filogenias de distintos grupos de organismos nos proporciona hipótesis acerca de la historia evolutiva y también de los procesos que la han generado.
Déjenme ofrecerles dos usos de la filogenia: el primer ejemplo tiene que ver con establecer cuáles especies están más cercanamente emparentadas en un grupo; es decir cuáles de ellas son “especies hermanas”. Debido a que existe un reloj molecular en el cambio en una secuencia de adn, de tal manera que la diferencia en la misma secuencia entre dos especies hermanas se incrementa de manera más o menos constante a través del tiempo desde el momento en que se separaron, es posible calibrar la tasa de evolución de la secuencia, y esto nos permite, por primera vez, fechar la especiación, es decir establecer cuándo ocurrió la divergencia. La figura 1 muestra un ejemplo interesante. Nancy Nowells, quien fuera mi estudiante, investigó la especiación en el género Melanoplus de la familia Acrididae (Ortóptera). Este es un grupo monofilético de una especie de chapulines que habita en las Montañas Rocallosas en altitudes elevadas. Mi alumna se interesó en el patrón e historia de especiación y encontró varios resultados interesantes. Uno relevante es que la divergencia en la secuencia del adn mitocondrial de estas especies es de menos de 4%. Debido a que existen numerosas calibraciones para insectos en la divergencia en la secuencia de adn mitocondrial, su conclusión es que el grupo completo de especies se originó durante el Pleistoceno, esto es durante los últimos 1.8 millones de años. Como puede apreciarse en la figura 1, han ocurrido cuatro o cinco eventos sucesivos de especiación en un periodo relativamente corto, lo que indica claramente que el proceso de especiación ha sido extremadamente rápido, quizás como consecuencia de la fragmentación de poblaciones debido al avance de los glaciares durante el Pleistoceno.
|
||
 figura 1
|
||
| El segundo ejemplo tiene que ver con la especiación en aves. Cuando estudiaba mi doctorado se aceptaba que las especies de aves de Norteamérica cercanamente emparentadas habían especiado durante el periodo Pleistocénico. Sin embargo esta idea ha sido desafiada recientemente por investigadores como Klicka y Zink. Este último estudió distintos pares de especies hermanas de aves en Estados Unidos y analizó el grado de divergencia entre secuencias de adn. En la figura 2 se muestra el número de especies hermanas de aves comparado con la divergencia en la secuencia entre cada par. Uno esperaría que mientras más antigua sea la separación de dos especies hermanas mayor será la cantidad de divergencia entre ellas. Sin embargo, la conclusión de Klicka y Zink, reforzada por los datos presentados en la figura, indica que la mayoría de los pares de especies hermanas de aves que se pensaba que habían evolucionado recientemente en realidad divergieron durante el Plioceno (entre cinco y dos millones años), de manera que probablemente los eventos de especiación no fueron promovidos por el avance de los glaciares. De esta forma, los ejemplos anteriores nos indican que debemos pensar nuevamente la escala de tiempo a la cual la fauna moderna ha evolucionado. | ||
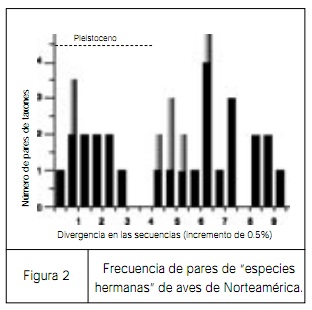 figura 2
|
||
|
Otro aspecto interesante de los estudios filogenéticos es que ahora podemos decir algo acerca de los mecanismos de la especiación. Por ejemplo, los estudios filogenéticos nos permiten describir la polaridad, es decir la dirección de la evolución de una característica. Si una característica tuviera dos estados en un grupo de especies emparentadas, nosotros podemos decir con frecuencia cuál de los estados es el ancestral y cuál es el estado apomórfico o derivado. Hasta hace poco era difícil afirmar esto. Hoy día además es posible decir si un estado derivado ha evolucionado por convergencia o por paralelismo.
Las especies de parásitos o de insectos herbívoros que pueden alimentarse de muchos hospederos se denominan generalistas, mientras que otras especies de parásitos o de insectos herbívoros que se alimentan de sólo una o de unas pocas especies de hospederos se denominan especialistas. Tradicionalmente se ha postulado que la especialización es un “callejón sin salida” evolutivo. Es decir que las especies especialistas no podrían evolucionar para convertirse en cualquier otra cosa y quedan “estancadas” en un nicho ecológico hasta que finalmente se extinguen. En resumen, la noción clásica es que las especies especialistas han derivado o han evolucionado a partir de especies generalistas, y que las especialistas no pueden originar especies generalistas.
Brian Farrell y su estudiante Kelley son de los primeros que han intentado abordar esta pregunta analizando un grupo de escarabajos del género Dendroctonus. Estos escarabajos atacan principalmente especies de coníferas; algunas especies están muy especializadas mientras que otras pueden alimentarse de distintas especies de coníferas. En la figura 3 es posible apreciar la filogenia del género Dendroctonus. Las especies generalistas de Dendroctonus están ilustradas en blanco y las especialistas en negro. Puede apreciarse claramente que las especialistas están siempre al final de las ramas del árbol filogenético, lo cual apoya la hipótesis tradicional de que a partir de las especies generalistas evolucionan las especialistas. Lo contrario no se aprecia en la filogenia del género Dendroctonus, y ésta es la primera vez que hemos sido capaces de poner a prueba la hipótesis tradicional sobre la evolución de los especialistas y generalistas. Esta pregunta es relevante porque está en la frontera de la ecología y la evolución. Sin embargo, después del análisis de Farrell, otros estudios como el de Nosil, en otros grupos de insectos han demostrado que la hipótesis tradicional no siempre es verdadera. Existen algunos grupos de especialistas que pueden dar origen a especies generalistas y algunas veces el fenómeno ocurre en ambas direcciones. De manera que, hasta el momento, no podemos concluir acerca de la evolución del generalismo y el “especialismo” ecológico, pero es probable que el estudio de otros grupos de parásitos haga posible llegar a una conclusión más sólida.
|
||
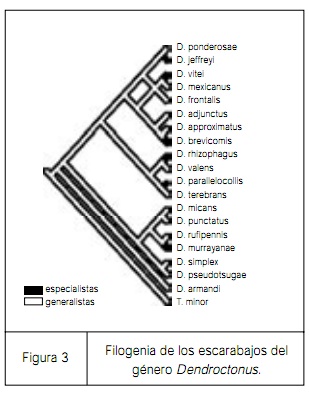 figura 3
|
||
|
La teoría de la coalescencia
El segundo tema importante que me gustaría mencionar es el estudio de lo que llamamos genealogía de genes, es decir filogenias de diferentes secuencias de adn de un locus entre especies o dentro de un grupo de especies cercanamente emparentadas. De manera que podamos trazar un ancestro común entre dos o tres especies, y podamos tomar tres secuencias variables de adn y, por los mismos métodos filogenéticos, llegar a la conclusión de cuáles secuencias de adn dentro de un grupo de especies son las más cercanamente relacionadas. Este hecho, en conjunto con la teoría matemática, es la base de la teoría de la coalescencia. La teoría de la coalescencia ve a la variación genética dentro de una población con una historia, o sea que los diferentes alelos o secuencias de adn dentro de una población tienen una historia, y que en última instancia todas las diferentes secuencias debieron de haberse originado de una secuencia original en el pasado. La mayoría de las secuencias pudieron haberse eliminado por deriva génica o quizás por selección natural, de manera que las secuencias existentes vienen de una secuencia ancestral y podríamos preguntarnos: ¿cuál es la secuencia ancestral?, ¿es reciente o es muy antigua? Y si pudiéramos tener esa estimación de la edad, entonces podríamos hacer inferencias acerca de la selección sobre esa secuencia, ya sea selección balanceadora o selección direccional. Algunas veces incluso es posible estimar el tamaño histórico de la población simplemente por el hecho de que una población que ha sido de tamaño muy grande, preservará las distintas secuencias por más tiempo, debido a que éstas no se extinguen rápidamente por deriva génica. También es posible, algunas veces, realizar una estimación de si la población ha crecido en tamaño o ha permanecido del mismo tamaño a través del tiempo.
Otro aspecto a estudiar de las genealogías de genes es la distribución geográfica de las diferentes variantes de un gen, que puede ofrecer una estimación de la historia de los cambios en la distribución de las poblaciones que constituyen la especie. Este es el campo de la filogeografía, un campo muy activo en el pasado reciente y que promete aportar una gran cantidad de información a la biología evolutiva. Veamos dos ejemplos.
Los genes de autoincompatibilidad en plantas evitan que un genotipo pueda producir descendencia por autopolinización. El árbol ilustrado en la figura 4 corresponde a distintos haplotipos, es decir, diferentes secuencias de un locus que produce autoincompatibilidad en las plantas de la familia Solanaceae. Lo que se observa en la figura 4 son diferentes formas o alelos del mismo gen, y puede apreciarse que en Nicotiana existen diferentes alelos de autoincompatibilidad. Hay cuatro diferentes alelos en este locus, y por lo tanto se dice que es un locus polimórfico, lo mismo puede decirse para el género Petunia. Pero lo relevante de este ejemplo es que los alelos del género Nicotiana (nic) están más cercanamente relacionados, no con otros del mismo género, sino con otros alelos que se encuentran en Solanum (sol), Petunia (pet) o Lycopersicon. Lo que significa que se trata de un polimorfismo muy antiguo, del cual ha habido linajes de genes por mucho tiempo y que todos, o varios de estos linajes de genes, fueron heredados por Nicotiana, Petunia o Solanum. En resumen, quiere decir que este polimorfismo ha existido desde el ancestro común de todos estos géneros de Solanaceae. Otra pregunta que podríamos hacernos es: ¿Qué tan antiguo es o por cuánto tiempo ha persistido este polimorfismo? La respuesta no la sabemos con certeza, pero el origen podría remontarse al Mioceno, de 30 a 40 millones de años. Y esto significa que probablemente ha estado bajo selección balanceadora, es decir una selección que ha mantenido el polimorfismo en la población. Hoy, por ejemplo, sabemos que opera la selección dependiente de la frecuencia sobre los alelos de autoincompatibilidad en plantas; sin embargo, lo que no sabíamos es lo poderosa que puede ser la selección para mantener un polimorfismo por 30 millones de años o más. Éste es un hallazgo sorprendente.
|
||
 figura 4
|
||
| El segundo ejemplo acerca del tipo de inferencias que podemos obtener de una filogenia de genes está relacionado con nosotros mismos, los seres humanos. La figura 5 muestra un árbol filogenético derivado a partir de la variación en secuencias de adn mitocondrial en humanos y en chimpancés. En el árbol tenemos una secuencia para chimpancés y varias para humanos de diferentes partes del mundo, de donde provienen estas secuencias. Lo primero que observamos es que todas las secuencias antiguas están presentes en África, y la longitud de las ramas indica la cantidad de diferencias mutacionales que existen entre las secuencias. Los resultados también indican que las secuencias son muy antiguas, o sea que han permanecido por mucho tiempo, el suficiente para acumular diferentes mutaciones, y por lo tanto indican claramente que las secuencias más antiguas existieron en África y después se extendieron al resto del mundo. De manera que tenemos evidencia incontrovertible de que África fue el hogar original de Homo sapiens, lo que por supuesto ya conocíamos por el registro fósil; en segundo lugar sabemos que los humanos han habitado África por mucho más tiempo que ninguna otra parte del mundo y que en ese continente los humanos han tenido una población más grande que en el resto del mundo. También podemos apreciar, gracias al adn mitocondrial, que las secuencias de otras partes del mundo son casi idénticas. Han ocurrido muy pocas diferencias mutacionales de un lugar a otro, como se aprecia por lo corto de las ramas. De manera que todas las secuencias mitocondriales debieron haber descendido de una secuencia ancestral común en el pasado reciente y posteriormente de grupos poblacionales pequeños que colonizaron el resto del mundo. Este aspecto es uno de los más interesantes que se han descubierto acerca de la historia evolutiva de nuestra especie. Esta aproximación también puede ser empleada para el estudio de otras especies. | ||
 figura 5
|
||
| Otro aspecto derivado de la teoría de la coalescencia y que se relaciona con el tamaño poblacional ancestral y la retención de variantes o de secuencias de genes, se puede apreciar en la figura 6a, que muestra una genealogía de genes relacionados entre sí y que difieren sólo en cambios mutacionales, pero todos derivan de un ancestro común. Cuando la población sea muy grande, todos los linajes de genes serán retenidos en la población por largo tiempo y tendrán muchos genes que estarán muy cercanamente relacionados, por lo que sólo existirán pequeñas diferencias entre ellos. Mientras más diferencias existan entre dos secuencias, más lejano en el pasado estará el ancestro común. Pero, debido a que la población es de tamaño grande, estos linajes de genes se mantendrán por mucho tiempo. De manera que si tomamos pares de secuencias de adn para este locus y calculamos el porcentaje de pares que tienen cero, una, dos, tres o más diferencias, esperaríamos una distribución de frecuencias como la que se ilustra en la figura 6a. Sin embargo, si la población ha sido pequeña y ha pasado por “cuellos de botella” poblacionales, es probable que los genes que existen hoy día vengan de un ancestro común reciente. Si la población está en expansión acumulará diferencias mutacionales, pero debido a que no ha habido suficiente tiempo para crear un árbol completo, la distribución de las frecuencias de las diferencias entre pares de genes será totalmente distinta a aquella derivada de poblaciones de tamaño grande (figura 6b). | ||
 figura 6
|
||
|
Por supuesto, si aplicamos esto a las poblaciones de humanos en Asia y en Europa, ambas aparecen como si hubieran sido originadas por poblaciones pequeñas.
Dicha teoría se puede aplicar a la evolución de nuevas especies. Uno de los postulados acerca de cómo surgen las especies fue hecho por Ernst Mayr en 1954, denominado teoría de la especiación por efecto del fundador. Mayr postuló que una nueva especie frecuentemente tiene su origen en poblaciones pequeñas, fundadas por pocos individuos. Si esto es verdad, será posible apreciar en las especies así originadas la “firma” de un cuello de botella en la variación en las secuencias de adn. Precisamente esto fue lo que estudiamos en el escarabajo Ophraella. En la filogenia de este escarabajo, figura 7, tenemos a las especies Ophraella communa, ampliamente distribuida desde el sur de Canadá hasta Guatemala, y Ophraella bilineata, distribuida únicamente en el borde de la distribución de la primera. Ambas están muy cercanamente emparentadas y supusimos que quizás Ophraella bilineata se originó en el borde de la distribución, justo como Ernst Mayr postuló para el origen de muchas especies nuevas. Nos preguntamos si la especie Ophraella bilineata mostraría una reducción en la variación genética y una distribución única de las frecuencias de las secuencias. Los resultados indican claramente que la población periférica no muestra evidencia de haberse originado por efecto de un fundador, y por lo tanto no apoyan la hipótesis de Ernst Mayr sobre el origen de nuevas especies a partir de un pequeño número de individuos. De manera que la teoría de la coalescencia puede usarse para explorar algunos aspectos de la especiación. Si una población ha sido fundada por unos cuantos individuos, uno esperaría que tuviera un número pequeño de secuencias diferentes, secuencias de diferentes haplotipos que probablemente estarían cercanamente relacionados uno con otro y que difirieran sólo en unas cuantas mutaciones. Aquí es donde entra la filogeografía, es decir la distribución geográfica de una secuencia de genes en relación a la filogenia de los genes.
|
||
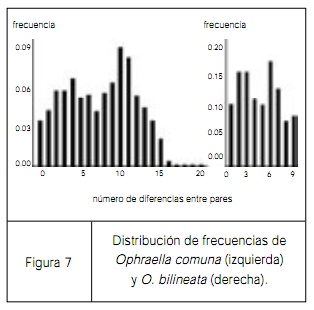 figura 7
|
||
|
La filogeografía
El siguiente ejemplo es acerca de un ave pequeña, Oporornis tolmiei, cuya distribución geográfica se presenta en la figura 8. Esta ave tiene una población reproductiva en un área enorme del oeste de Estados Unidos y en Canadá. En la figura también se muestra otra área en el este de Coahuila y oeste de Nuevo León; un área mucho más pequeña de una población disyunta con respecto a la costa oeste de Norteamérica. Se analizaron tanto la población mexicana como la de Estados Unidos, usando secuencias de adn mitocondrial, y se construyó el árbol de genes y de las localidades de las cuales se tomaron las muestras. Los resultados indican que hay un haplotipo que es muy común en los Estados Unidos y está presente en casi todas las poblaciones al igual que en México. Mientras que este haplotipo es el más común y es casi el único haplotipo en Estados Unidos, aunque tiene algunas variantes que difieren por mutaciones únicas o sencillas, las muestras tomadas de la población reproductiva en México son mucho más variables en las secuencias de adn mitocondrial; aquí existen múltiples haplotipos que son igualmente frecuentes y que tienen hasta cuatro diferencias mutacionales.
|
||
 figura 8
|
||
|
Esto nos indica que este grupo de haplotipos es mucho más antiguo y ha persistido por mucho más tiempo en México que en Estados Unidos. La distribución de frecuencias de las diferencias en las secuencias de la población de los Estados Unidos es unimodal, lo cual sugiere que esta población creció recientemente a partir de un número pequeño de individuos. Por el contrario, la distribución de frecuencias de la población mexicana tiene una forma que sugiere que la población original ha sido grande. ¿Qué quiere decir ésto? Esto sugiere que la población de esta ave en México es la población ancestral y que probablemente en la actualidad es una población relicto en esta área geográfica de México. Esta evidencia indica que la historia de las especies y sus distribución geográfica pueden ser inferidas a partir de árboles de genes.
La especiación es muy difícil de estudiar debido a que usualmente es un proceso sumamente lento para ser observado, pero también es temporalmente muy corto como para quedar grabado en el registro fósil. Por ello, casi cada aspecto de la especiación es controvertido. Pero hoy existe la esperanza de poder responder a las viejas preguntas acerca de la especiación. Por ejemplo, hoy poseemos nuevas herramientas para analizar las diferencias genéticas entre especies, de manera que podemos abordar el análisis del aislamiento reproductivo entre especies, su aislamiento conductual, mecánico o fisiológico y saber si producen híbridos y éstos son estériles. Hoy también podemos preguntarnos si el aislamiento reproductivo es producido por una diferencia de uno, dos o tres genes, o si ésta requiere que haya diferencias en muchos más genes. Intentamos conocer cuánto tiempo toma el proceso de especiación. Tenemos indicios, como los de los estudios de los chapulines y de las aves, de que el proceso de especiación a veces puede ser muy rápido.
También podemos preguntarnos si la especiación simpátrica es posible, es decir aquella que ocurre cuando no hay barreras en el flujo genético entre las poblaciones que ocupan una misma área. También he mostrado que podemos preguntarnos si la especiación por el efecto fundador es probable o no. Podemos tener alguna guía acerca de qué tan frecuentemente surgen las especies por el efecto fundador, justo como lo sugirió Ernst Mayr, pero una pregunta que podríamos abordar y que casi nadie ha contestado sino hasta muy recientemente es la siguiente: ¿Es la especiación producida por selección natural? La mayoría de los evolucionistas ha supuesto siempre que el aislamiento reproductivo es producido por la selección natural. Una idea popular entre los evolucionistas, sugerida por Ernst Mayr y Theodosius Dobzhansky, es que las poblaciones geográficas de una especie se adaptan a diferentes condiciones ecológicas y que los cambios genéticos tendrán efectos pleiotrópicos, es decir efectos colaterales que causan alteraciones en el comportamiento, desarrollo o fisiología, y que las hacen incompatibles con otras poblaciones, por lo cual no pueden intercambiar genes, ni se cruzan o son incapaces de producir descendencia viable y fértil. En resumen, la idea es que la especiación es un producto secundario de la selección natural ecológica; la selección, al adaptar localmente a las poblaciones, favorece indirectamente el aislamiento reproductivo.
Pero si uno revisa los libros de texto, incluida la primera edición de mi libro Biología evolutiva, y busca ejemplos acerca del papel de la selección natural en el origen de nuevas especies o del aislamiento reproductivo, difícilmente encontrará uno debido a que nadie se había fijado en esa pregunta sino hasta hace muy poco. Aunque todo mundo asume que la selección natural produce especiación, el hecho es que no lo sabemos con certeza. ¿Es posible que dos poblaciones que están aisladas sufran cambios por deriva génica, es decir cambios puramente aleatorios, y se vuelvan los suficientemente distintas genéticamente, de manera que sean incompatibles una con otra? Esto es totalmente plausible. De hecho sabemos muy poco acerca de si la selección natural o la deriva génica son responsables de la especiación. Hasta hace poco sólo un pequeño número de investigadores había entendido lo importante que es esta pregunta: ¿Qué es lo que causa la evolución de nuevas especies? Es decir, ¿qué puede ser más importante que eso? Uno de mis estudiantes, Daniel Funk, intentó contestar la siguiente pregunta: ¿La adaptación a diferentes condiciones ecológicas acelera la evolución del aislamiento reproductivo? Para ello, estudió insectos herbívoros, especializados en diferentes especies de plantas, y se preguntó si en poblaciones de una especie adaptadas a diferentes especies de plantas hospederas, la adaptación divergente acelera el origen del aislamiento reproductivo entre aquellas poblaciones geográficas. Supongamos que tenemos dos poblaciones separadas y que ambas se alimentan de la misma planta A (la población 1 y la población 2). Es probable que exista cierta divergencia genética entre ambas poblaciones debido a distintas fuerzas evolutivas, algo de deriva génica, quizás diferencias producidas por el clima o por diferentes depredadores, es decir, divergencia genética causada por otras causas distintas al aislamiento reproductivo. Pero si tenemos dos poblaciones que ocupan diferentes plantas hospederas, entonces habrá las mismas fuerzas evolutivas misceláneas que dan origen a la divergencia genética, además de las diferencias que surgen por su adaptación a diferentes plantas hospederas. Y si la hipótesis de Mayr de que el aislamiento reproductivo evoluciona como un efecto colateral de la divergencia genética debido a las circunstancias ecológicas es cierta, entonces en las diferentes poblaciones asociadas a un hospedero distinto evolucionará más rápido el aislamiento reproductivo que en las poblaciones que ocupan el mismo hospedero. Para poner a prueba esta hipótesis, Funk eligió una especie de escarabajo crisomélido del género Neochlamisus, en particular la especie Neochlamisus bebbianae. Este escarabajo se alimenta de tres especies de plantas; en gran parte del este de Norteamérica así como en Ontario, Canadá, se alimenta del maple rojo (Acer rubrum). Hay otras poblaciones que por el contrario se alimentan de sauces (género Salix) y las de Oklahoma se alimentan del abedul (género Betula). Funk tomó poblaciones que se alimentaban del maple rojo en Nueva York y en Georgia como el par de poblaciones que se alimentan del mismo hospedero; también tomó poblaciones que se alimentaban de Salix y también de Betula. Lo que hizo entonces en el laboratorio fue poner parejas en cajas de Petri: macho y hembra provenientes del mismo o diferente hospedero y observó su comportamiento. Registró el porcentaje de parejas que copularon y produjeron huevos; por otra parte, también demostró que estas poblaciones de insectos están adaptadas a diferentes plantas hospederas, de manera que las poblaciones que se alimentan de Salix tienen una mejor adaptación a este hospedero que los escarabajos que se alimentan de Acer. Una medida del aislamiento sexual es la proporción de parejas que no se aparean. El primer resultado fue que las poblaciones de Georgia que se alimentan de maple no mostraron aislamiento sexual con las poblaciones de Nueva York, que también se alimentan de maple, es decir que no existe barrera reproductiva entre ellas. Pero cuando tomaba poblaciones de Georgia o de Nueva York que se alimentan de maple y las ponía junto al sexo opuesto de las poblaciones de Ontario que se alimentan de Salix, lo que encontró fue un alto grado de aislamiento reproductivo. Lo mismo ocurrió cuando comparó a los escarabajos que se alimentan de Acer con los que se alimentan de Betula: cuando puso un macho proveniente de Betula con una hembra de Acer o viceversa encontró un alto grado de aislamiento sexual. De manera que, tal como se predijo, el aislamiento sexual es más alto entre poblaciones que están adaptadas a diferentes plantas hospederas. Funk también analizó el adn mitocondrial debido a que las diferentes poblaciones de diferentes hospederas pudieran ser más viejas, y de esta manera la divergencia se explicaría por el tiempo durante el cual se han acumulado las diferencias genéticas por deriva génica. Pero lo que encontró fue que las dos poblaciones que se alimentan de Acer eran más viejas; es decir que tuvieron un antepasado común más antiguo que el de Salix de Canadá, y por lo tanto hay menos tiempo de divergencia entre la población de escarabajos que se alimenta de Salix que de la población que se alimenta de Acer en Nueva York. Los resultados de este estudio apoyan la hipótesis de Mayr y Dobzhansky sobre la especiación o el aislamiento reproductivo como un producto secundario de la adaptación ecológica a diferentes circunstancias; en este caso plantas hospederas.
Evolución de las interacciones ecológicas
¿Cómo interactúan las especies una con otra, ya sea depredador y presa, parásito y hospedero o mutualistas y competidores? ¿Cómo evolucionan en conjunto? Y es aún más compleja la pregunta, ¿cómo es que la estructura de una comunidad ecológica depende de la evolución?
Veamos brevemente algo acerca de la evolución del uso de recursos, ya que éste es un tópico muy importante para la interacción de especies y subyace en gran parte de la teoría de la ecología de las comunidades. Cuando estudiaba mi doctorado, Robert MacArthur y su mentora Evelyn Hutchinson eran los ecólogos más importantes y creían que era posible producir una teoría general de la estructura de las comunidades, basada parcialmente en la teoría de la evolución. La idea era que, por ejemplo, las especies que compiten evolucionarían para segregarse en lo que llamamos nichos ecológicos, de tal forma que habría una explotación consistente o una división de los recursos entre un cierto número de especies que minimizaría la cantidad de competencia entre ellas. La evidencia más clara de apoyo a esta idea proviene del trabajo de Jim Brown, quien mostró que en dos desiertos de Norteamérica las especies de roedores de diferentes tamaños, que se alimentan de semillas, en gran medida están segregadas en diferentes clases de recursos en relación al tamaño del roedor. La comparación entre las comunidades mostró que había un paralelismo, es decir, una correspondencia de especies en ambos desiertos, y esto fue tomado como el ejemplo de que la estructura de la comunidad puede predecirse y que puede evolucionar a una clase consistente, casi óptima, como una función de la evolución de estas especies con otras.
Pero una de las grandes preguntas, que considero aún permanece en ecología evolutiva, es si este cuadro optimista de la predecibilidad de las comunidades es verdadero o no. Y la respuesta parece ser que algunas veces lo es y otras no. Veamos lo que considero como el ejemplo más espectacular en apoyo a la teoría de Robert MacArthur: las lagartijas del género Anolis en las Antillas. En las grandes islas, Cuba, Puerto Rico, Haití y Dominicana, y en Jamaica, existen varias especies de lagartijas del género Anolis que se segregan en distintos hábitats, de manera que unas viven en las copas de los árboles, otras viven en el tronco o corteza, algunas en pequeñas ramas e incluso las hay que viven en el suelo o en la base de los árboles. Pero el hecho más notable es que en la filogenia de las especies de Anolis en cada una de las Grandes Antillas encontramos similitudes impresionantes: en primer lugar, el hecho ecológico es que las Anolis están segregadas en diferentes hábitats, pero la filogenia indica que en cada isla el grupo de Anolis es monofilético. De manera que esto implica que Anolis ha irradiado, en respuesta al mismo conjunto de circunstancias en el uso de recursos; este es un caso de radiación adaptativa, y es extraordinario porque muestra la predecibilidad de la evolución entre especies que compiten y se segregan en distintos nichos de manera consistente. Sin embargo este fenómeno no siempre ocurre. Por ejemplo, los escarabajos de la tribu Tetraopini (familia Cerambicidae, orden Coleoptera), cuyo origen se remonta al Oligoceno (35 millones de años), se alimentan sólo de plantas de la familia Apocynaceae, y esta restricción histórica de un grupo de insectos a un grupo particular de plantas indica que estos insectos no han sido capaces de adaptarse rápidamente a diferentes clases de hospederos. A diferencia de las especies de lagartijas Anolis, que pueden reaccionar a la presencia de competidores evolucionando un cambio en la forma en la cual usan los recursos disponibles, parece que muchos grupos de insectos no tuvieran esa capacidad para evolucionar hacia un cambio de su planta hospedera particular, de forma que pudieran expandirse a distintas especies de plantas en una comunidad. El resultado de esto es que en muchos grupos de insectos existe un impacto de la historia evolutiva a largo plazo en la estructura contemporánea de la comunidad de plantas y de insectos. Esto da como resultado que ciertas especies de plantas tengan menos clases de insectos alimentándose de ellas, mientras que otras especies de plantas tendrán muchos insectos. Y esto es, en parte, una cuestión que tiene que ver con la cantidad de tiempo evolutivo en el cual ha habido oportunidad para los insectos de adaptarse a las plantas cuando son nuevas para ellos. Por ejemplo, la familia de plantas Asteraceae tiene muchas tribus distintas; algunas de estas tribus llegaron a Norteamérica hace sólo unos cuantos millones de años y tienen una muy pequeña fauna de insectos alimentándose de ellas; sin embargo, otras tribus como Astereae y Heliantheae son mucho más viejas en Norteamérica y tienen un vasto número de especies asociadas. De manera que la estructura contemporánea de la cadena alimentaria depende en gran medida de qué tan antigua sea la historia evolutiva en el caso de las interacciones de planta e insecto, mientras que en el caso de las lagartijas del género Anolis ha ocurrido un acomodamiento evolutivo de una especie a otra.
Epílogo
En los párrafos precedentes he tratado de señalar algunas de las preguntas que se analizan en ciertas áreas de la biología evolutiva en las cuales estamos atestiguando un grado de progreso y una profundidad en el entendimiento que no habíamos alcanzado antes. Si miro a las ediciones anteriores de mi libro de texto, Biología evolutiva, y comparo lo que dice con lo que sabemos hoy, me digo, ¡por Dios!, ¿yo escribí esto? Es tan diferente de lo que sabemos hoy que es casi como otro siglo. Y este progreso tiene implicaciones importantes fuera del ámbito académico del estudio de la evolución debido a que los biólogos evolucionistas, como muchos otros biólogos, están entendiendo que tienen cosas importantes que decir que son de valor social para la comunidad como un todo. Tenemos muchas cosas que decir acerca de la evolución de los parásitos, incluidos los microbios y los patógenos; tenemos muchas cosas importantes que decir acerca del papel de los genes en el desarrollo y los efectos pleiotrópicos de los genes en la especiación. El análisis de los efectos genéticos sobre el fenotipo es importante si uno empieza a pensar sobre los organismos transgénicos o genéticamente modificados por humanos. Existen muchas formas en las cuales la biología evolutiva es aplicable a cuestiones sociales de nuestra era, desde la salud humana hasta la conservación de las especies que todos apreciamos. Por lo tanto, quiero reiterar que éste es uno de los tiempos más excitantes para ser un biólogo evolutivo y me congratulo de que muchos de quienes estén aquí escuchado lo sean.
|
||
|
Traducción
Juan Núñez Farfán
Conferencia presentada en la Facultad de Ciencias el 28 de noviembre de 2002.
Transcripción: Mónica Benítez.
|
||
|
Referencias bibliográficas
Funk D. J. “Isolating a role for natural selection in in speciation: host adaptation and sexual isolation in Neochalamisus Bibbianae haf Beattlis”, en Evolution, núm. 52, pp. 1744-1759.
Futuyma, D. J. 1979. Evolutionary biology. Sunderland, Massachussets.
Ingman, M., H. Kaessmann, S. Paabo y U. Gyllensten. 2000. "Mitochondrial genome variation and the origin of modern humans", en Nature, núm. 408, pp. 708-713.
Ioerger, T. R., A. G. Clark y T. H. Kao. 1990. "Polymorphism at the self-incompatibility locus in Solanaceae predates speciation", en Proceedings of the National Academy of Sciences, Estados Unidos, núm. 87, pp. 9732-9735.
Kelley, S. T. y B. D. Farrell. 1998. "Is specialization a dead end? The phylogeny of host use in Dendroctonus bark beetles (Scolytidae)", en Evolution, núm. 52, pp. 1731-1743.
Klicka, J., and R. M. Zink. 1997. "The importance of recent ice ages in speciation: a failed paradigm", en Science, núm. 277, pp. 1666-1669.
Knowles, L. L., D. J. Futuyma, W. F. Eanes y B. Rannala. 1999. “Insight into speciation from historical demography in the phytophagous beetle genus Ophraella”, en Evolution, núm. 53, pp. 1846-1856.
Knowles, L. L. 2000. “Tests of Pleistocene speciation in montane grasshoppers (genus Melanoplus) from the sky islands of western North America”, en Evolution, núm. 54, pp. 1337-1348.
Mila et al. 2000. “Genetic evidence for ten effect of a postglacial population expansion on the phylogeography of a Northamerican Sungbird”, en Proceedings of the Royal Society of London, núm. B 267, pp. 1033-1040.
Nosil, P. 2002. "Transition rates between specialization and generalization in phytophagous insects" en Evolution, núm. 56, pp. 1701-1706.
Figuras
1. Cortesía de L. L. Knowles; 2. Klicka y Zink, 1999; 3. Kelley y Farell, 1998; 4. Ioerger et. al. 1990; 5. Ingman et. al. 2000; 6. Cortesía de Ingman et. al. 2000; 7. Cortesía de L. L. Knowles, 8. Mila et. al. 2000.
|
||
|
Douglas J. Futuyma
Departamento de Ecología y Biología Evolutiva, Universidad de Michigan.
_______________________________________________________________
como citar este artículo → Futuyma, Douglas J. (2003). Principales líneas de estudio en la biología evolutiva contemporánea. Ciencias 71, julio-septiembre, 16-28. [En línea] |
||
| ←Regresar al índice / artículo siguiente→ | ||
 |
 |
|
| De grandes y pequeñas cosas o la paradoja de Banach y Tarski |
|
|
|
Laura Elena Morales Guerrero
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Porque todas las cosas, muy grandes o muy pequeñas, pueden traerse dentro del dominio humano con la ayuda de las matemáticas y de la imaginación. Pues a diferencia de los científicos, quienes observan la naturaleza con los cinco sentidos, los matemáticos lo hacen con el sentido de la imaginación. Es su sexto sentido y están tan especializados en su uso como lo están los músicos con los sonidos, los gourmets con los sabores y aromas, o los fotógrafos y los cineastas con el sentido de la vista. A través de sus creaciones únicas los matemáticos nos informan de la realidad sin pretender siquiera demostrar que algo existe o no. Uno de los orígenes de las matemáticas está en la naturaleza juguetona del ser humano y es por eso que las matemáticas no son sólo una ciencia sino también un arte. Las matemáticas son una creación de la mente, una colección de cosas que existe sólo en la mente, indistinguibles unas de otras, y una colección de declaraciones acerca de estas cosas que se toman por ciertas. Tales declaraciones relacionan estas cosas inventadas o imaginadas, y a partir de ellas el matemático descubre otras llamadas teoremas, las cuales se aboca a demostrar. En suma, el matemático es un artista, su medio es la mente y sus creaciones las ideas.
Las matemáticas son erróneamente consideradas como la ciencia del sentido común. Pero éstas trascienden el sentido común y van más allá de la imaginación y de la intuición. Esto no quiere decir que las matemáticas no tengan relevancia en las creencias y actividades irracionales características de los seres humanos, no. Entender las matemáticas nos puede ayudar enormemente en nuestros intentos multidisciplinarios por entender la naturaleza humana. Escribiremos aquí sobre matemáticas sin usar fórmulas y aun así podremos expresar algo de su sentir, aunque será como expresar el sentimiento de un soneto sin la forma de soneto. No obstante, algo del espíritu de las matemáticas se puede salvar. Tendría entonces que quedarse sin respuesta una pregunta como: ¿qué son las matemáticas? Cualquiera que sea su esencia, son tan libres como la mente y tan enganchadoras como la imaginación. Las matemáticas son el propio trabajo del ser humano sujeto solamente a las limitaciones impuestas por las leyes del pensamiento. La matemática es una actividad gobernada por las mismas reglas impuestas a las sinfonías de Beethoven, a las pinturas de Da Vinci y a la poesía de Homero. De la misma forma que las escalas musicales, las leyes de la perspectiva y las reglas de la métrica parecen no tener lustre, las reglas formales de las matemáticas pudieran parecer opacas. Pero las matemáticas alcanzan pináculos tan altos como los logrados por la imaginación en sus artistas más atrevidos. Esto encierra, quizás, la paradoja última de la ciencia: el mundo de la razón pura es más extraño que el mundo de la pura fantasía.
El espacio matemático
Todo lo que tenga que ver con lo infinitamente pequeño o lo infinitamente grande es paradójico. En aritmética aprendemos que el todo es mayor que sus partes. Pero al tratar con el infinito matemático lo primero que encontramos es una paradoja: el todo no es mayor que ninguna de sus partes. Qué no habría dicho Zenón de esto, él, tan escéptico que era acerca de lo obvio. Las matemáticas de lo infinitamente grande deben mucho a Georg Cantor. En cambio Karl Weierstrass estuvo muy ocupado disponiendo de lo infinitesimal. De la existencia de lo infinitamente pequeño podría reírse cualquiera pero, ¿quién se atrevería a hacerlo de lo infinitamente grande? Ciertamente, Cantor, no. El estudio del infinito en las matemáticas de los números puros es un reto a la intuición, a nuestras experiencias cotidianas y debe ser respetado como tal. Por otro lado, en la vida real, entre las convicciones más preciadas ninguna como nuestras creencias acerca del espacio y tiempo (¿es el espacio infinitamente grande?), pero ninguna más difícil de explicar. Más aún, una cosa es el espacio físico, el de la percepción sensorial, y otro el del espacio matemático. Para entender este último se deben dejar de lado todas las nociones preconcebidas y aprender de nuevo el alfabeto. Hablar del espacio matemático quiere decir hablar de geometría, de varias clases de geometría. Los matemáticos las crean, y no precisamente en relación con figuras. Estas geometrías no tratan con nada real; no describen el espacio accesible a nuestros sentidos, el que explicamos en términos de ver y tocar. Hablan acerca de puntos que no tienen dimensiones, de líneas que no tienen anchura y de planos sin espesor. Ninguna de las abstracciones e idealizaciones se parece a nada de lo que hemos encontrado o experimentado. Más aún, estos espacios no se restringen a las tres dimensiones en las que vivimos (o en las que creemos vivir), sino que pueden ser de 4, 5, …n dimensiones. Y si estas nuevas geometrías son útiles para algo, eso no le concierne al matemático. Él es el sastre de la ciencia: hace los trajes, cualquiera que quepa en ellos puede usarlos. Para ponerlo de otra forma, el matemático hace las reglas del juego; el que quiera puede jugar en tanto observe esas reglas. No tiene sentido quejarse después de que el juego no sirvió para nada.
Por ejemplo, la noción de una cuarta dimensión, aun cuando es muy precisa, es muy abstracta; para la gran mayoría está más allá de la imaginación. El desarrollo de estas ideas obedece más a un deseo infantil de consistencia que a cualquier otra cosa. Con esas mismas motivaciones de consistencia y generalidad los matemáticos desarrollaron los números negativos, los números imaginarios y los trascendentales. No fue sino a través de una lucha que estas ideas, ahora comunes, se introdujeron en matemáticas, ya que nadie ha visto menos tres vacas o la raíz cuadrada de menos un árbol. Algo similar sucede con la cuarta dimensión. Comenzando, como es usual, con Aristóteles, se probó una y otra vez que una cuarta dimensión era impensable e imposible; pareció concluir por inducción que no hay transferencia en otra geometría y el admirable Tolomeo lo probó. No obstante, la geometría de cuatro y aun dimensiones mayores es una parte indispensable de las matemáticas. Pero eso no es todo. Por alguna razón misteriosa de las que siempre hay, felizmente, las aplicaciones de la geometría de cuatro dimensiones a la física, al mundo físico, trajeron al mundo el niño no deseado: ¡el tiempo!, que fue entonces reconocido y bautizado como la cuarta dimensión. Los físicos podrán considerar al tiempo como una cuarta dimensión pero el matemático no. Él se queja de su incapacidad para mostrar la cuarta dimensión como algo más concreto que el tiempo.
Una primera paradoja
Entre las innovaciones en matemáticas del último cuarto de siglo está el desarrollo de un par de teorías: la de conjuntos puntuales y la de funciones de variable real. Basándose enteramente en estos métodos de análisis matemático se logró un mayor rigor y generalidad en la geometría; mayor de lo que podría imaginarse en el caso de que la ciencia se hubiera desarrollado sólo por medios intuitivos. Se encontró que todas las ideas geométricas convencionales, incluyendo la idea de la “geometría de hule” (la topología), se podrían redefinir con exactitud mayor apelando a la idea de agregados y a las nuevas herramientas de análisis. Sin embargo una paradoja notable que se mantuvo escondida es suficiente, en sí misma, para mostrar que nuestras ideas intuitivas acerca de dimensionalidad y área, no carentes de precisión, son a menudo, completamente confusas y que no bastan las nuevas herramientas para resolverlas. Esta paradoja es una construcción puramente teórica de conjuntos matemáticos. Las piezas involucradas en una partición así son tan diferentes, dentadas, recortadas en los bordes de un modo desigual y tan exóticas, que no tienen una noción de volumen bien definida o una medida asociada a ellas. Así, una pelota que tiene un volumen definido se puede dividir en muchas piezas que se pueden reensamblar, por rotaciones en el espacio real, para formar dos o aun un millón de pelotas ¡cada una idéntica a la original! La forma de las piezas desafía nuestra concepción de área y volumen. Lo que la paradoja muestra es que no importa qué tanto se esfuerce nadie en definir un volumen, lo cual parece muy intuitivo para nosotros, de manera que corresponda con nuestra definición usual para conjuntos “bonitos”, siempre habrá conjuntos “malos” para los cuales sea imposible definir un volumen.
La paradoja surge al tratar de asegurar que se puede asignar un número, llamado medida, a cada figura en el plano para que se satisfagan las tres condiciones siguientes: 1) dos figuras congruentes tienen la misma medida. La palabra congruente se usará en el mismo sentido en que se aprendió en geometría elemental, es decir, en el sentido de que dos figuras son congruentes si ocupan el mismo lugar en el espacio aun en posiciones distintas; 2) si una figura se divide en dos partes, la suma de las medidas asignadas a cada una de las dos partes es exactamente igual a la medida asignada a la figura original; 3) como modelo para determinar el método de asignar una medida a cada figura en el plano, se acordó que la medida 1 debería asignarse al cuadrado cuyo lado tiene longitud 1. Este concepto de medida equivale al área de una figura en un plano. Pero debe tenerse en cuenta que este concepto se introdujo como un ejercicio general y teórico y no como la vasta y obviamente imposible tarea de realmente medir cada figura concebible. El problema se consideraba resuelto si se podía dar una prueba teórica de que a cada figura se le podría asignar una medida única; el enfoque debía ser analítico (por medio de conjuntos puntuales) y no geométrico. Aquí viene entonces la debacle. Se descubrió el hecho sorprendente de que el mismo problema, cuando se extendía a las superficies, no sólo no era soluble sino que llevaba a las paradojas más desconcertantes. En el caso de la superficie de la esfera, por ejemplo, los mismos métodos que resultaron fructíferos al investigar en el plano fueron inadecuados para determinar una medida única. Las condiciones para asignar una medida a una superficie son similares a las de las figuras en el plano dadas arriba, con la salvedad de que la condición 3 cambia a: si S denota la superficie total de una esfera de radio r, la medida asignada a S será 4πr2. El avance en la teoría de funciones y los nuevos métodos de análisis resolvieron algunos de estos problemas, pero introdujeron otros relacionados con el infinito, y la presencia de ese concepto, como todos los matemáticos lo han sabido desde siempre, de ningún modo es una bendición. Han podido avanzar a grandes saltos hacia delante, sí, pero no sin la sombra de la incertidumbre. Por supuesto que se puede uno quedar con la fórmula para el área de la esfera por la simple razón de que funciona. Pero si alguien se quiere mantener al paso del abierto e incansable espíritu matemático se topa con las inconfortables alternativas de abandonar la lógica para mantener los conceptos clásicos o aceptar los resultados paradójicos del nuevo análisis y mandar a volar lo sensato.
La paradoja de Hausdorff
El matemático alemán Hausdorff construyó una paradoja en verdad notable y mostró que el problema de la esfera es insoluble, que no se puede asignar una medida única a la superficie de la esfera de forma que las condiciones mencionadas se satisfagan. Mostró que si la superficie de una esfera se divide en tres partes distintas separadas: A, B y C, de manera que A sea congruente con B, y B sea congruente con C, surge una paradoja extraña que nos recuerda fuertemente algunas de las paradojas de la aritmética transfinita, y de hecho está relacionada con ellas. Hausdorff probó que no sólo A es congruente con C, como se esperaría, sino que también A es congruente con B+C. ¿Cuáles son las implicaciones de este resultado desconcertante? Veamos; si se asigna una medida a A, la misma medida debe asignarse a B y a C, dado que A es congruente con B, B es congruente con C, y A es congruente con C. Pero, por otro lado, puesto que A es congruente con B+C, la medida asignada a A tendría que ser la misma que la suma de las medidas asignadas a B y a C. Obviamente tal relación podría valer sólo en el caso en que las medidas asignadas a A, B y C fueran cero. Pero eso es imposible por la condición 3. De acuerdo con ella, la suma de las medidas asignadas a las partes de la superficie de una esfera debe ser igual a 4πr2. ¿Cómo entonces se asigna la medida? Estamos ante una contradicción desesperanzada.
La paradoja de Banach y Tarski
Dos distinguidos matemáticos polacos, Banach y Tarski, extendieron las implicaciones del teorema paradójico de Hausdorff al espacio tridimensional con resultados tan increíbles y alarmantes que no encuentran igual en todas las matemáticas. Las conclusiones, aun cuando son rigurosas e impecables, son casi tan increíbles para el matemático como para el lego. Imaginemos dos cuerpos en el espacio tridimensional: uno muy pequeño, como un chícharo, y otro muy grande, como el Sol. Pensemos también en una naranja entera. Recordemos que nos referiremos no a las superficies de estas esferas sino a las esferas sólidas completas. La paradoja de Banach y Tarski sostiene que en teoría es posible cortar una naranja en cierto número de piezas que pueden reensamblarse para producir dos naranjas, cada una ¡del mismo tamaño y volumen que la original! La versión alterna dice que es posible cortar un chícharo en un número finito de partes y reensamblarlas para formar una pelota sólida ¡del tamaño del Sol!
Para tener una idea de cómo se hace esto en la teoría, llamemos S al Sol y dividámoslo en muchas partes pequeñas. Cada parte es independiente y distinta de las demás y la totalidad de las partes es un cierto número. Desígnense estas partes con s1, s2, s3,… sn, que juntas hacen la esfera total S. Similarmente el chícharo será S’ y se dividirá en un número igual de partes mutuamente excluyentes, s’1, s’2, s’3,… s’n, las cuales juntas forman el chícharo. La proposición dice que si el Sol y el chícharo se han cortado de tal forma que la pequeña porción s1 del Sol es congruente con la partecita s’1 del chícharo y s2 es congruente con s’2, y s3 lo es con s’3 y así sucesivamente hasta que sn sea congruente con s’n, el proceso agotará no sólo todas las partes del chícharo sino también todas las partes del Sol. En otras palabras, el Sol y el chícharo se han dividido de manera que un cierto número de partes disyuntas de cada uno es congruente con una única parte del otro y después de que cada pequeña porción del chícharo se ha comparado con una pequeña porción del Sol no sobra ninguna parte del Sol. Esto es una simple correspondencia uno a uno entre los elementos del conjunto que corresponde al Sol y los elementos del otro conjunto que corresponde al chícharo. La paradoja está en el hecho de que cada elemento es comparado con otro con el que es completamente congruente o idéntico en forma y tamaño y que hay suficientes elementos en el conjunto que hace al chícharo para comparar con los elementos que hacen al Sol. Sí, suena terrible, descabellado. ¿Cómo es posible que haya una forma de dividir una esfera tan grande como el Sol en partes separadas donde ningún par de partes tenga ningún punto en común y aun así, sin comprimir o distorsionar ninguna parte, el Sol entero pueda ajustarse de manera que podamos sostenerlo en la palma de la mano? Más aún, las partes componentes del chícharo pueden reensamblarse, sin expandirse o distorsionarse y sin que ninguna tenga ningún punto en común con otra, de forma que llenen el universo entero sin dejar ningún espacio libre, sea en el interior del chícharo o en el Universo mismo. Solamente con herramientas matemáticas especializadas podemos explicar cómo una aseveración tan en contra de la intuición puede ser cierta.
Ningún cuento de hadas, ninguna fantasía de noches árabes ni sueño afiebrado alguno pueden compararse con este teorema de dura lógica matemática. Aun cuando los teoremas de Hausdorff, de Banach y Tarski, no pueden, hasta el momento, ser llevados a la práctica ni siquiera por aquellos que esperan poder empacar sus abultadas pertenencias en un pequeño maletín de fin de semana o por quienes esperan poder reproducir los huevos de oro, permanecen como un reto magnífico para la imaginación y como un tributo a la concepción matemática. Porque únicamente la imaginación pone un límite a las aplicaciones del teorema de Banach y Tarski.
Pero la paradoja no es ningún sin sentido. Debido al axioma de elección, existen conjuntos que no tienen un volumen medible. Es posible entonces descomponer los conjuntos y reensamblarlos con un volumen mayor. Esto no funciona si no se cree en el axioma de elección. El cual es precisamente eso: un axioma. Es decir que se ha probado que el axioma de elección no puede ser probado o desaprobado y es motivo de discusiones técnicas y filosóficas su uso en el teorema de Banach y Tarski porque en la prueba de este teorema se usó el axioma de elección. Y es posible mostrar que es imposible hacerlo sin el axioma de elección. Seguramente Banach y Tarski se sintieron como Einstein cuando quiso deshacerse del principio de incertidumbre en física. Al parecer Banach y Tarski esperaban que su resultado animara a los matemáticos a descartar el axioma de elección. Se sintieron infelices cuando la respuesta general fue: ¡Hey, el axioma de elección es fabuloso!, ¿de qué otra forma podríamos obtener resultados “contra-intuitivos” tan bonitos? Se ha argumentado que el resultado es tan “contra-intuitivo”, tan patentemente falso en el mundo real, que una de las suposiciones fundamentales debe ser incorrecta, y generalmente se escoge al axioma de elección como el culpable. Sin embargo, resultados extraños que no dependen del axioma de elección abundan en matemáticas. Por otro lado, el axioma de elección es consistente con los otros axiomas de la teoría de conjuntos. De ahí que la paradoja de Banach y Tarski sea, al menos, consistente.
Las falacias matemáticas
Hay otros juegos mentales que no son propiamente paradojas y se conocen como falacias matemáticas. Surgen tanto en aritmética como en geometría y se encuentran, aun cuando no muy a menudo, en las ramas altas de las matemáticas como, por ejemplo, en cálculo o en las series infinitas. Además de su aspecto divertido, muestran cómo una cadena de razonamientos puede viciarse por completo debido a un paso mal dado, aunque la mayoría de ellas son demasiado triviales para merecer atención.
Un ejemplo famoso trata del infinito. El infinito, en matemáticas, es siempre ingrato, a menos que sea tratado propiamente. Una serie que preocupó a Leibnitz es la terriblemente simple suma de unos: 1-1+1-1+1-1 +1…ad infinitum. Apareando los términos de maneras distintas, se obtiene una variedad de resultados; por ejemplo, (1-1)+(1-1|)+(1-1)+…=0. Pero también, 1-(1-1)-(1-1)…=¡1!
Al igual que en los cuentos y leyendas populares, las paradojas lógicas tienen sus antecedentes en los tiempos antiguos. Habiéndose ocupado de la filosofía y de las bases de la lógica, los griegos formularon algunos acertijos y problemas capciosos que en tiempos recientes han vuelto a contaminar a los matemáticos y a los filósofos. Muchos de ellos, no resueltos con el uso de la lógica, son causados por lo que se conoce como la falacia del círculo vicioso, la cual se origina al “ignorar el principio fundamental de que lo que involucra el todo de un total dado, no puede, en sí mismo, ser un miembro de la totalidad”. Ejemplos simples de esto son esas frases pontificias que parecen tener un gran significado pero que en realidad no tienen ninguno. Tales como: “Nunca digas nunca”. “Todas las reglas tienen excepciones”. “Todas las generalidades son falsas”.
Entre algunas de las más avanzadas está la siguiente: “El barbero del pueblo rasura a todos los que no se rasuran a sí mismos”. Este principio pronto lo envuelve en una contradicción dialéctica: ¿debe el barbero rasurarse a sí mismo? Si lo hace entonces está rasurando a alguien que se rasura a sí mismo y rompe la propia regla. Si no lo hace, además de quedarse barbudo, también rompe su regla al no rasurar a una persona del pueblo que no se rasura a sí misma… El uso de la palabra todos es peligroso en matemáticas.
En otra situación, considérese el hecho de que cada número entero puede expresarse en palabras, sin usar símbolos. Así se tiene que 1 400 se puede escribir mil cuatrocientos o 1 769 823 como un millón setecientos sesenta y nueve mil ochocientos veintitrés. Es evidente que ciertos números requieren más palabras que otros; en general, mientras más grande sea el número, se necesitarán más palabras para expresarlo. El primer caso requiere dos palabras, el segundo nueve. Ahora bien, puede establecerse, ¿por qué no?, que ciertos números requerirán más de doce palabras y otras menos de doce. Más aún, no es difícil mostrar que entre los números que requieren exactamente doce palabras hay uno que es el más pequeño de ellos. Entonces, es fácil ver que “el número más pequeño no expresable en menos de doce palabras” es una frase que describe al número específico 101 131 131 (ciento un millones ciento treinta y un mil ciento treinta y uno) en ¡11 palabras! Por tanto tenemos una contradicción: el número menor expresable en doce palabras, se puede expresar también en las once palabras de la frase entre comillas.
Los matemáticos y las paradojas
Y así como estos casos contradictorios, hay otros en matemáticas. Tal vez la paradoja más grande sea que haya paradojas en matemáticas. Las paradojas lógicas aquí presentadas no son trucos vanos o tontos, no fueron presentados para hacer reír al lector a menos que su risa sea causada por las limitaciones de la lógica. Parafraseando a Hamlet “lo que una vez fue paradoja no lo es más pero podría volver a serlo”. La matemática moderna, en un intento por prevenir las paradojas, se enfrentó abiertamente con la alternativa de adoptar un escepticismo aniquilador con respecto a todo el razonamiento matemático o reconstruir tanto los fundamentos de las matemáticas como los de la lógica. Mucho han trabajado los matemáticos y los filósofos en este sentido sin haber podido llegar a una solución. Lo que se mantiene es el hecho de que las paradojas lógicas han dividido a los matemáticos en facciones opuestas e irreconciliables y tienen que ser eliminadas. Se ha enfatizado el hecho de que el matemático busca siempre trabajar sus teoremas en la forma más general posible (inclusive cuando no puede resolver un problema, ¡lo generaliza!), en un reclamo de mucha altura. Ese pasar del uno al todo también es peligroso; peligroso en la misma forma en que el concepto de infinito lo es. En la transición de lo particular a lo general el matemático ha hecho sus más grandes progresos, pero también ha sufrido sus más grandes reveses. De entre ellos las paradojas lógicas constituyen la parte más importante.
|
||
|
Laura Elena Morales Guerrero
Instituto de Matemáticas, Universidad Nacional Autónoma de México.
_______________________________________________________________
como citar este artículo → Morales Guerrero, Laura Elena. (2003). De grandes y pequeñas cosas o la paradoja de Banach y Tarski. Ciencias 71, julio-septiembre, 46-54. [En línea] |
||
| ←Regresar al índice / artículo siguiente→ | ||
 |
 |
|
| Genética de poblaciones, una perspectiva histórica |
|
|
|
Manuel Feria Ortiz y Adrián Nieto Montes de Oca
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Desde principios del siglo XX han aparecido varios trabajos que consideran diferentes aspectos relacionados con la genética de poblaciones. Alrededor de 1930, R. A. Fisher y J. B. S. Haldane desarrollaron modelos para predecir el destino de alelos mutantes sujetos a selección. Estos trabajos, y en particular otro de S. Wright escrito por las mismas fechas, proporcionaron los principales elementos teóricos de la genética de poblaciones. Algunos años más tarde, T. Dobzhansky difundió en términos más sencillos las conclusiones principales de la genética de poblaciones. La labor que realizó, en conjunción con la de otros evolucionistas tales como E. Mayr y G. G. Simpson, ha permitido que los principios teóricos de la genética de poblaciones puedan aplicarse a otras áreas de la biología. Sin embargo, desde un inicio existieron posiciones opuestas dentro de esta disciplina biológica: algunos autores, como Dobzhansky y Dubinin, pensaron que los organismos deberían ser heterocigotos en la mayoría de sus loci (punto de vista equilibrado), mientras que otros, como Haldane, T. H. Morgan, y H. J. Muller, supusieron que la variabilidad genética era mínima y que la mayoría de los organismos deberían de ser homocigotos en casi todos sus loci (punto de vista clásico). A principio de la década de 1960, la aplicación de la técnica de electroforesis que hace posible el análisis del adn, reveló que las poblaciones naturales comúnmente exhiben una alta variabilidad genética, lo cual comprobó el punto de vista equilibrado. No obstante, las diferencias siguieron existiendo, pero ahora respecto al significado de la variabilidad genética. Para algunos, la mayor parte de ésta debería de ser adaptativamente neutra, y por lo tanto la deriva génica sería la fuerza evolutiva principal en el nivel molecular (punto de vista neutralista). Para otros, los diferentes alelos serían desde el punto de vista adaptativo más o menos adecuados y por lo tanto la selección natural positiva sería importante tanto en el nivel morfológico como en el molecular (punto de vista selectivo).
Los primeros trabajos que prepararon el camino para que posteriormente pudiera establecerse la genética de poblaciones aparecieron a finales del siglo XIX. En este tiempo se desarrolló un enfoque estadístico para el estudio de la herencia, mismo que fue comenzado y dirigido inicialmente por un primo de Charles Darwin, Sir Francis Galton. Más tarde fue dirigido por un discípulo de éste último, K. Pearson, auxiliado por R. F. Weldon. Estos “biometristas”, como ellos mismos se nombraban, estaban interesados en caracteres tales como el peso, la estatura y la inteligencia, los cuales varían de modo continuo de un individuo a otro. Sus métodos de análisis les permitieron percibir fácilmente que estas características por lo común se distribuyen de manera gausiana y que tal distribución sólo cambia ligeramente de una generación a la siguiente. En consecuencia, al igual que Darwin, concibieron la evolución como el cambio minúsculo y constante que experimenta la población como un todo. Así, a finales del siglo xix y principios del xx, los biometristas estuvieron entre los pocos naturalistas que apoyaron el esquema darwiniano de la evolución por selección natural.
Cuando se descubrió el escrito de Mendel en 1900, los biometristas se dieron cuenta de que la herencia de los rasgos que estudiaban no seguía las leyes establecidas por este autor. Asimismo se opusieron a las ideas de genetistas mendelianos como William Bateson, Hugo de Vries y Archival E. Garrod, quienes no sólo defendían la generalidad de los principios de Mendel sino que resaltaban la importancia de las macromutaciones en el cambio evolutivo de los organismos. De este modo comenzó una serie de discusiones entre mendelistas y biometristas que persistió durante casi tres décadas.
A principios del siglo xx la mayor parte de la investigación se centró en problemas de genética mendeliana. Por el contrario, el estudio de la evolución cayó en decadencia.
De manera paralela a los trabajos anteriores se desarrollaron otros conceptos que posteriormente serían fundamentales para el establecimiento de la genética de poblaciones. En 1902 Yule señaló que si en una población los individuos se aparean de manera aleatoria, las frecuencias genotípicas iniciales se mantienen en la población descendiente. Un año más tarde, el genetista americano W. E. Castle hizo referencia a las circunstancias bajo las cuales las frecuencias genotípicas de una población se mantendría constantes de una generación a otra, y cinco años después, de manera independiente, el matemático inglés G. H. Hardy y el médico alemán W. Weinberg, establecieron formalmente las condiciones bajo las cuales la composición genética de una población permanecería constante generación tras generación.
En un artículo teórico titulado “On the Correlation Between Relatives on the Suppositions of Mendelian Inheritance”, elaborado en 1918, el estadista inglés Ronald Aylmer Fisher demostró que los resultados obtenidos por los biometristas eran compatibles con los principios mendelianos. Su trabajo fue particularmente importante en la integración de ambas perspectivas, lo cual permitió eliminar las diferencias entre ellas.
También desde principios del siglo pasado, algunos trabajos contribuyeron al conocimiento de otros aspectos de la genética poblaciones. Por ejemplo, W. E. Castle realizó experimentos para conocer los efectos de la endogamia en la mosca de la fruta Drosophila melanogaster. También con el fin de comprender los efectos de la endogamia, Sewall Wrigth realizó experimentos con conejillos de indias y posteriormente, en 1922, definió algunos conceptos importantes tales como el coeficiente de endogamia, lo cual hizo posible generalizar la regla de Hardy-Weinberg a situaciones distintas a la del apareamiento aleatorio. En 1918 Sumer abordó otro aspecto al analizar las diferencias genéticas que existen entre las subespecies de Peromiscus maniculatus.
La genética de poblaciones
La integración de las escuelas biometrista y mendeliana y la valoración del efecto de la endogamia en las poblaciones constituyeron un paso fundamental en el desarrollo del enfoque matemático de la genética de poblaciones. El siguiente paso consistió en analizar el efecto de las diferentes fuerzas evolutivas, en particular la selección natural, en la constitución genética de las poblaciones. Este paso se llevó a cabo en gran parte gracias a los trabajos de R. A. Fisher, J. B. S. Haldane y S. Wrigth.
Fisher analizó el efecto de la selección natural en poblaciones grandes. Construyó un modelo para calcular la velocidad con la que un gen ventajoso aumenta su frecuencia en una población generación tras generación. Para Fisher, la selección es un proceso determinístico que actuaba de manera lenta pero segura, aumentando la frecuencia de genes favorables. De este modo, aquellos genes que resulten ventajosos se irán integrando al genoma, lo que conduciría a incrementar cada vez más la adecuación de los organismos. Fisher también analizó el caso de un sistema en el cual el heterocigoto tiene una ventaja adaptativa en comparación con los homocigotos y encontró que bajo esta situación los alelos involucrados permanecen en la población. Sus ideas quedaron plasmadas en un trabajo ahora clásico intitulado Genetical Theory of Natural Selection, publicado en 1930.
En 1932 Haldane publicó The Causes of Evolution, en el cual, al igual que Fisher, construyó un modelo matemático para analizar el destino de genes individuales sujetos a la selección natural. Haldane enfatizó la rapidez con la cual puede actuar la selección natural, tomando como ejemplo el cambio de coloración de la polilla Biston betularia en los bosques contaminados de Gran Bretaña. Estimó que a mediados del siglo xix, cuando las emanaciones de las industrias aún no contaminaban los bosques y los árboles estaban tapizados de líquenes blancos, la forma oscura de la polilla debería haber tenido una frecuencia extremadamente baja, mantenida por el balance entre su aparición por mutación y su eliminación por selección natural. No obstante, a partir de entonces los desechos industriales tendieron a oscurecer los troncos de los árboles y provocaron que las polillas oscuras se detectaran menos fácilmente que las claras cuando se posaban sobre este sustrato. En consecuencia, la frecuencia de la variante oscura comenzó a aumentar de tal modo que para 1900 casi había reemplazado a la variante clara, otrora predominante en las áreas boscosas de Inglaterra.
Si bien las contribuciones de Fisher y Haldane fueron muy importantes para el desarrollo de la genética de poblaciones, sus modelos sólo tomaban en cuenta el caso de poblaciones muy grandes con apareamiento aleatorio. En consecuencia, el cambio evolutivo se percibió como un cambio unidireccional, sin ramificaciones, y no dio oportunidad de que se establecieran asociaciones con otros fenómenos evolutivos importantes tales como la especiación.
En 1932 Wright adoptó un enfoque diferente. En lugar de concebir una población grande, consideró que una especie está integrada por una serie de poblaciones locales pequeñas (demos) en donde la endogamia, a pesar de ser limitada, es capaz de crear nuevos sistemas de interacción genética. Dentro de este contexto, los sistemas que resultaran ventajosos a los organismos serían favorecidos por la selección natural. Wright ideó un panorama genético, su paisaje adaptativo, compuesto por cimas de adecuación con diferentes alturas separadas por valles, áreas de baja adaptación, de profundidad también variable. En su modelo, los cambios aleatorios en la constitución genética de las poblaciones locales, o deriva génica, podrían provocar que una población local “descendiera” de una cima y se ubicara en la base de otra cima. En esta nueva situación, la selección natural conduciría a la población hacia un nuevo pico local, el cual podría ser más alto. De esta manera, bajo los efectos conjuntos de la selección, la deriva génica y la endogamia, una población podría cambiar de una cima adaptativa a otra.
Con las contribuciones de Fisher, Haldane y, en particular, la de Wright, la genética de poblaciones adquirió todos sus elementos básicos. No obstante, quedó planteada en términos eminentemente teóricos y, dada la complejidad de los modelos matemáticos involucrados, muy pocos biólogos apreciaron el alcance de la teoría. Además, los escritos de los fundadores de la genética de poblaciones prácticamente fueron desconocidos por aquellos naturalistas de campo que también estuvieron interesados en problemas evolutivos pero que abordaron el tema desde una perspectiva diferente. En consecuencia, la extensión y aplicación de la genética de poblaciones hacia otros campos de la biología tuvo que esperar un tiempo más.
El impacto de la genética de poblaciones
El evolucionista ruso Theodosius Dobzhansky revisó los trabajos teóricos de los genetistas de poblaciones, en particular los de S. Wright, y los planteó en términos más sencillos. Además, desde que llegó a Estados Unidos, en 1927, comenzó a trabajar extensivamente con poblaciones silvestres y cultivadas de moscas del género Drosophila. Los datos que obtuvo le permitieron, entre otras cosas, contrastar los modelos teóricos de la genética de poblaciones con la situación real encontrada en la naturaleza. De este modo, la labor de Dobzhansky creó un puente entre los planteamientos teóricos de los primeros genetistas y los resultados obtenidos por los biólogos de campo.
La obra principal de Dobzhansky, Genetics and the Origin of Species, se publicó por primera vez en 1937. La importancia que dio Dobzhansky a la genética de poblaciones se refleja en su afirmación siguiente: “dado que la evolución es un cambio en la composición genética de las poblaciones, los mecanismos de la evolución son asunto de la genética de poblaciones”.
El trabajo del biólogo ruso Sergei Chetverikov también jugó un papel fundamental en promover que la genética de poblaciones pudiera aplicarse en otros campos de la biología evolutiva. Chetverikov, al igual que Dobzhansky, trabajó con Droshophila. Mediante cruzas entre poblaciones silvestres demostró la existencia de una gran variabilidad genética al interior de las poblaciones, la cual normalmente no se puede apreciar debido a que están involucrados muchos alelos recesivos. Chetverikov fue darwinista e influyó de manera notable en el pensamiento de Dobzhansky.
Si bien autores como Fisher, Haldane, Wrigth y Dobzhansky ya habían abordado el tema en torno a la formación de nuevas especies, fue el ornitólogo Ernst Mayr, quién a través de su trabajo clásico Systematics and the Origin of Species, publicado en 1942, demostró más formalmente que este proceso podía explicarse con base en los principios teóricos de la genética de poblaciones. De acuerdo con este autor, los cambios que normalmente ocurren en la estructura genética de poblaciones geográficamente aisladas, dado el tiempo suficiente, pueden conducir a la formación de nuevas especies.
De este modo, gracias a los trabajos clásicos de autores como Dobzhansky, Mayr y Simpson, los principios teóricos de la genética de poblaciones pudieron aplicarse en otras áreas de la biología, lo que originó, en términos de T. H. Huxley, “la síntesis moderna de la teoría evolutiva”. Desde entonces, la genética de poblaciones quedó constituida como la base teórica fundamental de la biología evolutiva.
La estructura genética de las poblaciones
Por el tiempo en el que se integrara la síntesis hubo un aspecto que suscitó posiciones encontradas dentro de la genética de poblaciones. Éste consistió en tratar de establecer cuál debería de ser la naturaleza de la variabilidad genética en las poblaciones naturales, así como de los mecanismos responsables de mantener una estructura genética particular. Desde principios de la década de 1930, Haldane y T. H. Morgan, y después H. J. Muller, contribuyeron al desarrollo del denominado punto de vista clásico, el cual concebía a las poblaciones como líneas genéticas prácticamente puras. Partían de la suposición de que cada individuo debe ser homocigoto para un alelo silvestre o “normal” en la gran mayoría de sus loci. El estado heterocigoto existente en algunos pocos loci se debe a mutaciones, las cuales casi siempre son deletéreas y por lo tanto tarde o temprano son eliminadas por la selección natural. El número de generaciones necesarias para eliminar a un gen depende de la medida en que éste disminuye la adecuación del individuo. Dentro del contexto de la visión clásica, el papel de la selección natural consiste principalmente en eliminar o en mantener en frecuencias muy bajas a las mutaciones deletéreas que tienden a disminuir la adaptación de los organismos. En consecuencia, la variabilidad genética está directamente relacionada con la adecuación promedio de las poblaciones.
En 1950 Muller se refirió más explícitamente a la presencia de alelos deletéreos originados por mutación como una “carga de mutaciones”. Este concepto, denominado ahora simplemente carga genética, fue formalmente definido por Crow, en 1958, como: “la proporción en la que la aptitud (o cualquier otra característica que se considere) resulta disminuida en comparación con la de un genotipo óptimo”. Enfatizó que la carga genética puede originarse no sólo a través de la mutación, sino también a través de otros procesos como la segregación de homocigotos en un sistema en el que los heterocigotos tienen una mayor adecuación.
En oposición al punto de vista clásico, algunos autores, como Chetverikov y N. P. Dubinin, ya habían reparado en la existencia de muchos alelos deletéreos recesivos protegidos de la selección en individuos heterocigotos. No obstante, fue Dobzhansky el que a través de experimentos de cruza demostró más claramente la existencia de una gran cantidad de variación genética en poblaciones naturales de Drosophila. En contraposición con el modelo clásico, Dobzhansky propuso que la condición típica de un individuo particular es que sea heterocigoto en la gran mayoría de sus loci. Desde su punto de vista, la selección natural equilibradora es el principal mecanismo que mantiene los polimorfismos. Esta visión constituyó el modelo equilibrado de la estructura genética de las poblaciones, en el cual se reconoce la diversidad genética como un fenómeno fundamental en la naturaleza.
En 1966 Lewontin y Hubby utilizaron la técnica de electroforesis en gel para cuantificar la variabilidad genética en varias poblaciones de Drosophila pseudobscura y para mediados de la década de 1970 ya se había estudiado con esta técnica la composición genética de muchas especies de vertebrados e invertebrados. De este modo quedó demostrado que las poblaciones naturales típicamente poseen una cantidad enorme de variabilidad genética. Indudablemente, estos estudios dieron un apoyo considerable al modelo equilibrado y desacreditaron el punto de vista clásico. A pesar de esto, aún permanecía el problema de esclarecer el significado biológico de la variación. Una respuesta consistió en suponer que las diferentes variantes de algún gen o de una proteína particular poseían diferentes valores de adecuación, y por lo tanto su evolución debería de estar controlada principalmente por una selección natural dirigida.
No obstante, el genetista japonés M. Kimura propuso una respuesta diferente: se adhirió a la idea de que la mayoría de los nuevos alelos que aparecen por mutación son perjudiciales, pero consideró que una porción significativa de los mismos deberían de ser selectivamente neutros y que sólo unos cuantos resultarían ser ventajosos. Así, dada la rareza de las mutaciones benéficas y la predominancia de las deletéreas, la función principal de la selección consistiría en eliminar las mutaciones perjudiciales, mientras que el destino de las mutaciones neutras, las cuales explican la variabilidad genética presente en las poblaciones naturales, dependería de la deriva génica.
La teoría neutral de la evolución molecular
En 1971 Kimura y Otha propusieron que la variabilidad genética es un fenómeno transitorio en la evolución molecular. Su ocurrencia representa el balance entre los nuevos alelos que ingresan a la población a través de la mutación y la pérdida de los mismos por deriva génica. Argumentaron que la selección equilibradora no puede ser el mecanismo que mantiene el alto grado de polimorfismo observado en las poblaciones naturales, debido a que si esto fuera así, se generarían cargas genéticas absurdamente grandes.
Por otro lado, gracias a los avances de la biología molecular, para principios de la década de 1960 ya se había logrado obtener la secuencia de aminoácidos de las moléculas de la hemoglobina y del citocromo c en una variedad de organismos pertenecientes a distintos taxones. De nuevo, Kimura interpretó estos datos a la luz de la genética de poblaciones; con base en la comparación de las secuencias de aminoácidos de ambas proteínas, calculó la tasa de evolución de estas últimas. En seguida, extrapoló sus resultados a todo el genoma y estimó una tasa de evolución de una sustitución génica cada dos o tres años.
Previamente, en 1957, a través de una serie de suposiciones teóricas, Haldane había calculado que la selección natural únicamente podría provocar una tasa de evolución máxima de una sustitución génica cada trescientas generaciones. Una velocidad más alta implicaría un costo de selección, muertes genéticas necesarias para que ocurra la sustitución de un alelo por otro, demasiado alto como para permitir la existencia de cualquier especie. No obstante, la estimación de Kimura era casi cien veces mayor a este valor. Por ello, éste pensó que la selección natural no era el mecanismo que producía esta tasa de evolución. En cambio sugirió que a nivel molecular la evolución debería de estar gobernada principalmente por la deriva génica al actuar sobre mutaciones neutras o casi neutras.
Un año más tarde, con base en datos bioquímicos, King y Jukes llegaron a una conclusión similar a la de Kimura. Desde 1962, antes de que este último realizara sus estudios comparativos, Zuckerkandl y Pauling también habían comparado las secuencias de aminoácidos de las moléculas de la hemoglobina y el citocromo c en diferentes especies de mamíferos. Llegaron a un resultado diferente pero que tenía implicaciones importantes para la hipótesis propuesta por Kimura, King y Jukes: encontraron que en ambas moléculas la tasa de sustitución de aminoácidos era similar en diferentes linajes. Pensaron que este patrón de evolución era típico para todas las proteínas y propusieron lo que se conoce como la hipótesis del reloj molecular, de acuerdo con la cual la tasa de evolución de cualquier proteína es similar en los distintos linajes. En 1969 Kimura utilizó esta hipótesis para apoyar la suya propia respecto a la naturaleza de la evolución a nivel molecular.
Los estudios comparativos realizados por King y Jukes y por Dickerson durante la segunda mitad de la década de los sesentas y principios de los setentas, demostraron tres cosas: I) que las sustituciones de aminoácidos con propiedades fisicoquímicas similares son más frecuentes que las sustituciones entre aminoácidos con propiedades distintas; II) que las partes de las proteínas que son menos importantes para su función evolucionan más rápidamente que las partes que son más importantes; III) que las proteínas que realizan funciones más importantes para el organismo evolucionan más lentamente que las que realizan funciones menos relevantes. Estos hallazgos, de acuerdo con los neutralistas, apoyan la hipótesis de que en el nivel molecular la mayoría de las sustituciones alélicas ocurren entre variantes selectivamente neutras. Desde su punto de vista, las mutaciones que provocan el cambio de un aminoácido por otro equivalente o que afectan a partes o proteínas cuya función es poco relevante para el organismo, típicamente producen variantes proteicas con propiedades fisicoquímicas similares, que realizan su función con aproximadamente la misma eficiencia, y por lo tanto tienden a ser selectivamente neutras.
La controversia entre neutralistas y seleccionistas
Muchos autores rechazaron la hipótesis neutralista de la evolución molecular debido a que minimizaba el papel positivo de la selección natural y en consecuencia confinaba a la evolución adaptativa al nivel morfológico. De esta manera se originó una fuerte controversia entre los defensores de ambos puntos de vista, la cual actualmente es conocida como la controversia entre neutralistas y seleccionistas. Algunos autores, como Milkman y Maynard Smith, se opusieron al argumento de que si la selección equilibradora fuera el mecanismo responsable del polimorfismo detectado al interior de las poblaciones se generaría una alta carga genética. Según estos autores, si la selección favorece a un organismo particular puede favorecer simultáneamente a varios loci, de modo que si hay ventaja del heterocigoto en varios de ellos se puede favorecer una alta variabilidad genética sin generar cargas excesivas. Además, se señaló la existencia de otros modos de selección —como la selección dependiente de frecuencias—, los cuales pueden mantener la variabilidad genética sin necesidad de generar una carga genética.
Se originó una oposición similar a la idea de que la sustitución génica por selección natural no puede proceder a velocidades muy elevadas debido al alto costo en término de muertes que esto implica. Este razonamiento es aplicable sólo si se asume que ninguna de las muertes genéticas que se necesitan para que haya una sustitución génica están incluidas en las muertes que ocurren debido a las adversidades ecológicas; esto es si se asume que la selección es totalmente dura. No obstante, mientras las muertes selectivas estén representadas en la mortalidad ecológica natural, la selección natural podrá proceder a velocidades muy altas sin originar costos a la población. También se criticó la manera en que Haldane calculó el costo de selección, ya que, al igual que en el argumento de la carga genética, se consideró que los diferentes loci actúan de manera independiente. Aun así, entre más loci interactúen de manera dependiente menos realistas resultan las estimaciones del costo de selección.
Por otro lado, investigadores como Goodman y Czelusniak rechazaron la idea de que las proteínas evolucionan de manera constante. También hubo controversia respecto al efecto que debe tener el tiempo de generación sobre el ritmo de la evolución molecular; si la velocidad de sustitución de aminoácidos se debe a la deriva génica, como lo sugirió Kimura, entonces la evolución proteica debería reflejar el tiempo de generación: es decir que la velocidad de cambio debería ser mayor en organismos que tienen tiempos de generación cortos que en aquellos con tiempos de generación relativamente largos. No obstante, los datos muestran que una proteína dada evoluciona a una tasa similar en organismos con tiempos de generación largos o cortos.
A principios de los setentas Clarke sostuvo que el hecho de que la tasa de evolución de una proteína o parte de la misma dependa de la importancia de la función que realiza puede explicarse mejor desde un punto de vista seleccionista que desde uno neutralista. Las mutaciones con efectos estructurales grandes tienen mayores probabilidades de ser perjudiciales que las que provocan sólo cambios ligeros. Del mismo modo que las mutaciones que afectan a proteínas cuya función es poco relevante para el organismo tienen, con mayor probabilidad, efectos pequeños en el organismo, y es de esperarse que la tasa evolutiva sea más alta en este tipo de moléculas que en aquellas que realizan funciones fundamentales.
Simultáneamente, Otha modificó el modelo teórico de la teoría neutralista de la evolución molecular pues pensó que una porción grande de las nuevas mutaciones debería de consistir en mutaciones ligeramente desventajosas. La selección natural entonces debería actuar en contra de ellas. Sin embargo, dado el poco efecto que tienen sobre los organismos, su destino evolutivo dependería principalmente de la deriva génica. Con esta modificación, la teoría neutralista predice un nivel de heterosis menor al predicho si se piensa en mutaciones neutras. También bajo esta perspectiva, la tasa de evolución no tendría que ser constante y dependería del tamaño de las poblaciones; bajo neutralidad pura la tasa de evolución debe de ser igual a la tasa de mutación neutra. Así, esta modificación eliminó varias de las dificultades inherentes a la hipótesis neutralista pura.
En las décadas de los sesentas y setentas las posiciones teóricas en torno a la controversia entre neutralistas y seleccionistas se apoyaron principalmente en datos obtenidos en proteínas. No obstante, en las siguientes dos décadas, conforme el desarrollo de las técnicas de la ingeniería genética hizo posible la obtención de un número cada vez mayor de secuencias de adn y arn, la polémica dio un giro. Los cuatro hallazgos obtenidos con proteínas también aplican a nivel del adn: existe una gran cantidad de variabilidad genética (mucho mayor que en el caso de las proteínas), la tasa de evolución es relativamente alta y constante, y existe una restricción funcional. En consecuencia, muchos de los argumentos seleccionistas y neutralistas aplican tanto a las proteínas como al adn. No obstante, se ha observado que la teoría neutralista se ajusta más a los datos obtenidos con los ácidos nucleicos que a los obtenidos con proteínas. En el caso del adn se ha detectado que la tasa evolutiva refleja un efecto de tiempo de generación; esto significa que es más alta en organismos con tiempos de generación más cortos. Asimismo, se ha observado que la tasa de evolución de los pseudogenes es mayor que la de los genes funcionales. También, tal y como lo había predicho Kimura, la tasa de sustituciones silenciosas es mayor que la tasa de sustituciones significativas.
La controversia entre neutralistas y seleccionistas aún no termina. Pero ahora resulta claro que la deriva génica juega un papel mucho más importante del se había considerado en un principio. Desde otra perspectiva, también ha quedado demostrado que, al menos con lo que respecta a la evolución de las proteínas, el papel de la selección natural positiva no es tan insignificante como lo pensó Kimura.
|
||
|
Referencias bibliográficas
Ayala, F. J. (ed.). Molecular evolution. Sinauer Associates, Inc. Publishers, Ca., Massachussets.
Bowler, J. P. 1989. The mendelian revolution. The John Hopkins University Press, Baltimore.
Bowler, J. P. 1989. Evolution. The history of an idea. University of California Press, Berkeley.
Dobzhansky, T. 1970. Genetics of the evolutionary process. Columbia University Press, Nueva York.
Hanson, E. D. 1981. Understanding evolution. Oxford University Press, Nueva York.
Kimura, M. 1992. “The neutral theory”, en Fox K. E. y A. E. Lloyd (eds.), Key words in evolutionary biology. Harvard University Press, Cambridge, Ma.
Lenay, C. 1993. La evolución. De la bacteria al hombre. rba Editores, Barcelona.
Mayr, E. 1991. One long argument. Harvard University Press, Cambridge, Ma.
Panchen, A. 1993. Evolution. St. Marin’s Press, Nueva York.
Ridley, M. Evolution. Blackwell Scientific Publications, Oxford.
Ridley, M. (ed.). 1997. Evolution. Oxford University Press, Nueva York.
Rothhammer, F. 1977. Genética de poblaciones humanas. Secretaría General de la Organización de los Estados Americanos. Serie Biología, monografía Número 15.
Tamarin, H. R. 1996. Principios de Genética. 4a. ed., Editorial Reverté, Madrid.
Wright, S. 1960. “Genetics and twentieth century darwinism. A review and discussion”, en American Journal of Human Genetics, vol. 12, núm. 3, pp. 365-372.
|
||
|
Manuel Feria Ortiz
Facultad de Estudios Superiores Zaragoza,
Adrián Nieto
Facultad de Ciencias, Universidad Nacional Autónoma de México.
_______________________________________________________________
como citar este artículo → Feria Ortiz, Manuel y Nieto Montes de Oca, Adrián. (2003). Genética de poblaciones, una perspectiva histórica. Ciencias 71, julio-septiembre, 32-42. [En línea] |
||
| ←Regresar al índice / artículo siguiente→ | ||
 |
 |
|
| La divulgación de la ciencia en los tiempos de la postmodernidad |
|
|
|
Miguel Fernando Pacheco Muñoz
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Las acciones de comunicación y educación no formal sobre aspectos científicos se conocen en México como divulgación de la ciencia. Se le ha definido como disciplina, interdisciplina, filosofía práctica, arte, metodología y técnica social. La divulgación incluye implícitamente a la técnica y la tecnología, pero generalmente excluye de su discurso a las humanidades. Esto es una contradicción, ya que la divulgación cuenta con las características necesarias para ser considerada una disciplina de las ciencias sociales; en cierto sentido, la divulgación de la ciencia se da la espalda a sí misma, de tal manera que objetos de estudio y comunicación interdisciplinarios como la pobreza o el ambiente se muestran sumamente problemáticos, cuando no abiertamente despreciados y excluidos.
La divulgación es una práctica en expansión, en la que cada día hay nuevos actores; sujetos sociales e institucionales con nuevos conocimientos y diferentes saberes se suman al complejo campo de la divulgación. Así, al incorporarse traen consigo su agenda, sus experiencias y sus códigos profesionales. Todo lo cual permite que estos nuevos saberes influyan en el resto de la comunidad, ya sea integrándose o cuestionando las prácticas y fines ya establecidos.
De lo trivial o lo fundamental
Una visión que todavía se encuentra en la divulgación se caracteriza por utilizar un discurso centrado en los conocimientos. Esta corriente se basa en una noción de la ciencia que propone un culto o perspectiva casi mística del concepto de objetividad.
Se diferencia por una concepción de la ciencia libre de influencias ideológicas y políticas, en donde los sujetos, la sociedad y el mundo donde se desarrolla la ciencia no existen, no son importantes o están vedados.
Parece trivial decirlo, pero aquello que se divulga depende de lo que se considere o no como ciencia. Frente a una corriente objetivista y ahistórica del fenómeno científico se hace necesario replantear el significado de la divulgación científica dentro del contexto del llamado “giro histórico” o “nueva filosofía de la ciencia”. El objetivo de la articulación entre la divulgación y la nueva filosofía de la ciencia es ampliar los sentidos y significados de la divulgación. La carga semántica de una palabra que se utiliza para caracterizar un campo profesional no está delimitada sólo por su capacidad de designar con claridad y precisión una práctica, sino por las visiones ideológicas con las que se relaciona. Un claro ejemplo de esta situación la encontramos en el caso de los siguientes términos relacionados pero no idénticos: animación sociocultural, extensión cultural, difusión cultural. Como se menciona en el texto intitulado “Educación de adultos y educación popular”, cada grupo u organización, por su tradición, formación, origen, historia o posición ideológica, se siente más identificado con un término u otro para otorgar significado a lo que sus prácticas y trabajo constituyen.
Organizar una noción de divulgación a la que, en consecuencia con la nueva filosofía de la ciencia, pudiera llamarse “nueva divulgación” podría incluir una imagen de la ciencia que rebase las temáticas disciplinarias y una visión ahistórica de la ciencia; una divulgación que recupere los aportes de los estudios culturales de la ciencia, la filosofía, la sociología y la historia de la ciencia.
Así, la nueva divulgación de la ciencia estaría caracterizada por una visión sociohistórica del fenómeno científico, en la cual se analizan las relaciones entre la sociedad y la ciencia y viceversa.
Divulgación y educación, asociación o contradicción
La nueva divulgación puede ser empleada para abarcar tanto la forma comunicativa de la divulgación como su forma pedagógica. En toda forma comunicativa hay aspectos educativos y viceversa; educación y comunicación están indisolublemente ligadas. La divulgación es educación cuando no la reducimos a pedagogía y la comprendemos, en su sentido más amplio, como el proceso social por el cual aprehendemos y aprendemos la cultura. El rechazo a otorgar un papel educativo a la divulgación se basa en una concepción que confunde aprendizaje con enseñanza, o bien, limita lo educativo a la escuela.
En el debate sobre las diferencias entre educación y divulgación se encuentra el concepto de intencionalidad, éste ha sido uno de los criterios más utilizados para explicar la diferencia y se basa en el supuesto de que una actividad educativa intencional correspondería a las modalidades formales o no formales y, en contraparte, las que carecen de esta intencionalidad entrarían en el ámbito de la divulgación. Este argumento presenta debilidades, ya que, a pesar de que no se tenga la intención explícita de enseñar algo, siempre y, aunque no se quiera, se aprende algo implícitamente: una visión del mundo, una proposición sobre el ser humano o sobre la sociedad. Todas nuestras experiencias son educativas; no siempre aprendemos un contenido pero aprendemos sobre sentimientos, creencias o aptitudes hacia determinado fenómeno. No se requiere la existencia de un contrato educativo para aprender. Incluso su existencia no es garantía de aprendizaje o comprensión.
La divulgación había recurrido mayoritariamente a medios de comunicación, pero la estructura del campo cambió cuando se sumó la divulgación a través de los museos. Las prácticas clásicas como los materiales escritos, los programas de radio y las conferencias están siendo complementadas, integradas y hasta desplazadas, en cierto sentido, por la proliferación reciente de los museos de ciencia y las actividades educativas que giran en torno a ellos. La nueva divulgación tiene más posibilidades de incluir acciones de las diversas modalidades educativas. La formación de los departamentos de servicios educativos tuvo un impacto en el campo de la divulgación, creando nuevas propuestas y generando nuevas necesidades (teóricas, metodológicas, institucionales, etcétera).
La emergencia de esta relativamente nueva forma de divulgación, la educación no formal, tensiona ya los campos de los viejos divulgadores, sus formas y prácticas y las nuevas necesidades educativas en esta área. Ahora existe el peligro de formar una brecha entre comunicadores y pedagogos en un campo que apenas está curando las heridas de la escisión entre comunicadores y científicos.
Dentro o fuera de la caja de cristal
Otra razón que podemos argumentar en favor de una nueva divulgación es el rebasar la descripción o la naturaleza de las cosas. Superar un discurso que sólo esté basado en el contenido formal de la ciencia, en sus aspectos y postulados teóricos, sus leyes, teorías e hipótesis puede auxiliarnos a ir más allá de estos aspectos formales, para construir objetos de divulgación interdisciplinarios sobre problemas socialmente relevantes que se reflejen tanto en las producciones escritas como en la museografía, en los talleres y publicaciones.
No estamos pugnando por un pragmatismo en la selección cultural de la divulgación científica, al estilo de las propuestas de los discursos estatales sobre el financiamiento científico. La ciencia y el conocimiento que se obtiene de ella no están en una caja de cristal, aisladas de otras formas del conocimiento y la razón, de otros saberes; de la ética y la moral. La ciencia es cultura, de tal forma que es imposible escindir a la ciencia de la cultura, pues ella es cultura en sí misma, y tampoco es posible querer sustituir los valores por conocimientos.
He aquí otro aspecto que es combatido con el argumento de que la ciencia es ideológicamente neutra. Por ejemplo, los defensores de esta posición objetan que no ven ninguna idea política en un enunciado como: “La luz viaja en línea recta”. Tienen razón, no hay moral ni política en la naturaleza. Como bien lo explica Adolfo Sánchez Vázquez, no hay ciencias de derecha o izquierda, pero si la ciencia no es sólo la descripción de los objetos, entonces hay fines, intenciones y acciones sobre lo que se investiga, orientaciones de Estado, de mercado o de grupos sociales que plantean cómo se realiza la búsqueda del conocimiento, cómo se aplica y cómo se difunde. Todo esto se conoce como política científica y, como toda política, admite la distinción de derecha e izquierda o de algunas otras categorías analíticas que no tienen nada de neutrales. La divulgación es en sí misma un acto político, porque admite en ella la representación de valores, fines y creencias sobre todo lo relacionado con lo científico; en consecuencia el divulgador es un actor de la política científica. Los divulgadores necesariamente tendrían que pensar en los fines, valores y posiciones que asumen como actores de la política.
El momento de la ruptura
La nueva divulgación puede ayudar a aprehender los procesos de construcción, producción, distribución y apropiación de la ciencia y la tecnología. La ciencia es un cuerpo articulado de argumentos y explicaciones, pero además es las muchas maneras de llegar a ellos, por lo que se debería incluir en el discurso de divulgación la comprensión de las diversas posturas y escuelas de pensamiento sobre el conocimiento científico, es decir, la filosofía de la ciencia. Además, referirse a la evolución de las ideas y de las perspectivas sociales enmarcadas en un periodo y clase, a las posibilidades de aplicación técnica y a los espacios sociales y políticos para el desarrollo de una teoría, es pugnar por la inclusión de la historia de la ciencia. Incluir además los imaginarios sociales que sobre la ciencia y la técnica tienen las personas no es sólo un estímulo para la realización de una divulgación que utilize contextos históricos, económicos o sociales como notas de color, sino un intento por pensar y problematizar la divulgación desde la filosofía, la sociología y la historia de la ciencia.
A partir de la ruptura con el positivismo lógico del Círculo de Viena, de la publicación de La estructura de las revoluciones científicas de Thomas Kuhn y muy especialmente a partir de las polémicas entre Popper, Feyerabed y Lakatos, la filosofía de la ciencia ha cambiado el significado de la ciencia y su influencia se ha sentido en todos los campos que tienen como objeto de estudio el fenómeno científico. La divulgación no puede quedar al margen de esta influencia, pues si bien es cierto que la divulgación no estudia lo científico en sí, tal como lo hacen la filosofía y la sociología de la ciencia, la práctica de divulgación depende del propio concepto de ciencia que se maneje, de lo que se acceda a incluir o excluir.
Los estudios culturales de la ciencia no nos ofrecen una visión débil de la misma, la ciencia como producto humano está ligada indisolublemente a las personas, pues los científicos no son “objetos–objetivos”, sino que son sujetos. La ciencia no está manchada de internalidades subjetivas y externalidades de intercambio, sino que está constituida por y en ellos. No existe un conocimiento acultural y ahistórico; pensar así es una ilusión ideológica, un fantasma que confunde a los que se presumen dueños de lo racional, de los que creen que la ciencia los libera de su yo, del resto del mundo, de su responsabilidad, de las creencias y de la ideología.
Tal giro histórico es visto con gran desconfianza por algunos científicos temerosos de perder una razón querida y egoístamente atesorada. Algunos científicos ven a los historiadores y filósofos de la ciencia como nuevos enemigos que ponen junto a los dioses y los mitos y no tienen otros términos para ellos que irracionalistas y acientíficos. La posmodernidad resulta ser un terreno peligroso para la ciencia, y en muchos casos es posible que tengan razón, es decir, que haya propuestas en extremo radicales. Efectivamente, la ciencia no es sólo un juego del lenguaje o una fantasía lingüística, pero tampoco es el objeto inmaculado, ahistórico, apolítico, neutral, universal y unidireccional que insisten en presuponer.
El relativismo extremo es sólo una de las formas de la posmodernidad, pues no la ejemplifica toda. La posmodernidad no es homogénea, atiende a una gran diversidad de posiciones, algunas de ellas en evidente contradicción. La posmodernidad tiene muchos rostros y ésa es una de sus características, pero esas muchas caras pueden incluso ser contrarias. La posmodernidad es un conjunto heterogéneo de posturas filosóficas que se caracterizan por la crítica y el cuestionamiento a la modernidad, pero las coincidencias terminan ahí.
La posmodernidad puede ser entendida de varias formas: premodernidad, amodernidad, antimodernidad, transmodernidad, hipermodernidad, neomodernidad, todo ello con intereses transformistas, conservacionistas o reformistas. En el centro del debate entre la modernidad y la posmodernidad se encuentra uno de los elementos que definen lo moderno: la ciencia. La critica a la ciencia, a su proceso de conocimiento, a sus resultados, instituciones y personas tiene muchas facetas. De las diferentes escuelas posmodernas es desde donde se aprehende y crítica a la ciencia y su papel.
Los críticos de la posmodernidad ven sólo uno de los aspectos de esta confusión de principios de siglo; la condenan a muerte en nombre de la ciencia y la razón porque se extravían con las formas más radicales o porque no comprenden la ironía, la necesidad de leer entre líneas, para ver que decirle adiós a la razón no es una forma de decirle adiós a toda la razón, sino sólo a aquella que maximiza la ganancia y reduce las costos, a la que se olvida de los hombres y la naturaleza, a la razón fragmentada. Sin embargo parece ser que estamos condenados a los excesos; pasamos de un objetivismo extremo a un subjetivismo radical: creer que la ciencia va a salvar al mundo o creer que lo quiere destruir.
De razones y sinrazones
Son muchas las razones que se han expuesto para hacer divulgación: para compartir la pasión con que se realiza esta actividad, porque como espacio de la experiencia humana merece ser comunicada tal como se hace con las letras y las otras artes, por la necesidad de interesar a los jóvenes en formarse en este campo, para promover el aparato científico, la investigación, legitimar la labor del científico y mejorar la opinión pública sobre la agenda científica, para aumentar las capacidades productivas de la población, para que se adapten a los procesos técnicos y a la vida de un mundo globalizado, para incrementar la producción nacional o para incrementar la conciencia y la acción de las personas sobre los problemas del mundo.
En la divulgación el compromiso democrático resulta especialmente importante para que las personas tomen decisiones sobre lo que les afecta y se conviertan en verdaderos actores sociales e históricos; que no sean otros los que tomen las decisiones que afectan su vida. Si el ciudadano común no tiene el bagaje científico y cultural suficiente es fácil ser víctima de posiciones catastrofistas propias de mercaderes tecnológicos. Si no entendemos las complejas articulaciones ecológicas, económicas y políticas de la ciencia moderna es muy difícil proponer soluciones a los problemas del mundo contemporáneo. La falta de conocimiento se convierte en un instrumento de dominación, exclusión y marginación; sin éste, no es posible que las personas puedan tomar decisiones informadas sobre el futuro, que sean capaces de determinar y no sólo ser determinadas, capaces de construir una verdadera acción comunicativa. Para actuar en el mundo requerimos una plataforma cultural y un mínimo de conocimiento de los códigos del lenguaje, de la economía, la ciencia y el ambiente, mismos que nos permitan leer la realidad para construir el proceso democrático.
Una divulgación que ve a la ciencia como un producto de cultura de élite y pretende realizar un trabajo equiparable al de la difusión de las bellas artes, si bien puede tener las mejores intenciones del mundo, no hace más que llevar la ciencia a un nivel aristocrático mediante una visión excluyente, contraria a la visión democrática del saber y del poder. Como bien menciona Philippe Roqueplo: “El conocimiento en sí mismo, de una u otra forma, es un poder; resulta cuestionable que se pueda controlar ese poder sin apropiarse del propio conocimiento que le sirve de fundamento, el reparto del conocimiento tiene que plantearse el papel de las ciencias en el contexto político, social y económico que participa en la construcción y evolución tecnocrática de nuestras sociedades y por tanto la divulgación tiene un rol político”.
Hay que aclarar que esto no quiere decir que la divulgación no deba asegurarse de ser bella. La divulgación puede ser al mismo tiempo una tarea artística y una tarea política, un aspecto no está reñido con el otro. De hecho ya es una tarea artística y política, porque al elegir un fin ya se esta eligiendo una posición ideológica. El discurso de la divulgación propone, aunque el autor no se lo proponga, una serie de representaciones sociales, representaciones que finalmente también son morales. El significado está en el discurso, pero además está en lo que queda fuera de él, por lo que si sólo se plantea una posibilidad o ángulo de análisis, al ocultar o callar otros, estamos privilegiando una postura, un tipo de conocimiento o saber; lo que entra o sale del discurso, consciente o inconscientemente, es en sí mismo una posición ideológica, he ahí la ilusión de la neutralidad.
De esta forma, un discurso de divulgación centrado sólo en la belleza está promoviendo una visión apolítica y ahistórica de la ciencia, incluso puede estar promoviendo los intereses ya establecidos y legitimados, como dice Habermas. El discurso de la divulgación de la ciencia entendido como difusión cultural está basado en una pretendida pero inalcanzable neutralidad; cuando se trata de interesar por la ciencia sólo por su pasión o belleza el mensaje de neutralidad es altamente ideológico, pues se está enseñando sobre un sistema de pensamiento que enmascara las relaciones sociales que genera el conocimiento científico y este discurso por supuesto no es neutral sino ideológico.
Si la experiencia científica no es una práctica que opera en un vacío histórico, el discurso de la divulgación de la ciencia tampoco puede serlo. La divulgación es un conjunto de argumentos organizados que definen una agenda, es, per se, una narrativa que no sólo trata de comunicar un concepto científico sino que además lleva en ella una idea social que, como producto histórico en sí misma, surge de y se basa en la ciencia. Se trabaja en un contexto geográfico, cultural, espacial, temporal, profesional, etcétera. Así, se configura una concepción ideológica, positiva o negativa, sobre cierto aspecto de la realidad. Todos los textos y discursos, como intentos de comunicación, tienen un significado ético y no sólo estético. Una divulgación guiada por un esteticismo, una divulgación por el arte mismo, puede perfectamente ser calificada, paradójicamente para la divulgación, como neorromántica. Incluso, al convertir la estética en ética se acerca mucho a ciertas concepciones posmodernas que algunos divulgadores y científicos ven y citan como el “demonio antirracionalista”. La falta de compromiso social que hace ver a la divulgación como una forma de propaganda o producto mercadotécnico deja poco espacio para la reflexión sobre el mandato social de esta actividad y reduce sus posibilidades para compartir el conocimiento y el interés de la gente por apropiarse del mismo, se convierte en un arbitrario cultural que brinda la ilusión de un reparto democrático, cuando en realidad jamás llega a cuestionar la estructura jerárquica establecida para el reparto del conocimiento. Además, el discurso de la divulgación es parcial en el papel democrático de la ciencia en la sociedad, limitado para orientarla hacia metas y estrategias (construcción, aplicación y control) socialmente comprendidas y legitimadas. Puesto que su acción se circunscribe a la esfera superestructural, no puede encontrar en lo educativo la respuesta a todos los problemas. La ciencia y la técnica no tienen vida propia, como toda actividad humana están sujetas a las leyes de la historia; la ciencia y la tecnología son afectadas por las relaciones económicas: el conocimiento es una fuerza productiva. El cambio tecnológico permite generar nuevos productos, reducir los costos de producción, aumentar la productividad de la fuerza de trabajo y reducir el tiempo socialmente necesario para la reproducción del capital. Simultáneamente, la necesidad del capital por mejorar los procesos productivos ha impulsado el desarrollo de la ciencia y la tecnología. El desarrollo científico y tecnológico está íntimamente ligado al crecimiento económico y viceversa. Más sutilmente, como lo plantea Lyotard, como la ciencia de hoy está basada en la tecnología, ya que es a través de las mediaciones de los aparatos tecnológicos que se hacen las verificaciones, estos instrumentos no sólo implican conocimiento, sino dinero en la apropiación del conocimiento; así, los juegos del lenguaje científico se convierten en juegos donde el más rico tiene más oportunidades de tener razón.
Nos unimos a César Carrillo, para quien la divulgación de la ciencia debe dirigirse a reintegrar la separación disciplinaria, unir la ética al quehacer científico e integrar otras formas de conocimiento y saber. Pensamos, como él, que, al comunicar los límites y posibilidades de la ciencia y la tecnología, los procesos y estructuras sociales y económicos, los diversos principios filosóficos, epistemológicos, ideológicos y políticos que la condicionan, que determinan su producción, distribución y aplicación, en lugar de empobrecer a la ciencia y su divulgación, la enriquecen y permiten mostrar sus verdaderas posibilidades para la construcción de una sociedad democrática.
Algunos científicos ciegos a la pobreza, a la enfermedad, a la crisis ambiental, es decir, a la realidad, ven el discurso posmoderno como un ataque a la ciencia y presagian un nuevo oscurantismo, ven en la defensa de la ciencia la defensa de la verdad única y absoluta, cuando, muchas veces sin saberlo o entenderlo, lo que defienden implícita o explícitamente es el capital, las relaciones desiguales entre los hombres y la destrucción de la naturaleza. Por el contrario, una ciencia que no se cuestiona a sí misma nos llevará a un nuevo oscurantismo pero científico y tecnológico, pues si bien es cierto que el discurso de la posmodernidad critica a la modernidad y a la ciencia como su producto, no es para negarle su papel en el mundo. No le niega a la ciencia su papel en la historia, ni podría hacerlo, sino que trata de hacernos comprender que no es la única forma de conocimiento válido y legítimo; que el conocimiento científico no es condición directa del saber y no es la única solución a nuestros problemas.
Para los divulgadores de la ciencia la definición de nuestra actividad profesional pasa por una serie de discusiones acerca de su delimitación, y esto no es una cosa menor. No es sólo una confusión o un problema de énfasis, tipos o perspectivas; en el fondo esta discusión es un planteamiento de tipo ideológico, es una toma de postura, esencial para definir no sólo lo que es sino lo que nuestra práctica debe ser. Estos conflictos no son sólo diferentes aproximaciones teóricas o metodológicas a cierto problema, de hecho son una lucha entre diferentes visiones del mundo y, como dice Basil Berstein, son por lo tanto una batalla fundamentalmente moral.
|
||
|
Referencias bibliográficas
Bernstein, Basil. 1993. La estructura del discurso Pedagógico. Morata, Madrid.
Bacaicoa G., Fernando. 1988. Conflicto cognitivo. Universidad del País Vasco, Enseñar y Aprender.
Carrillo Trueba, César. 1997. “La divulgación de la ciencia en un mundo fragmentado”, en Ciencias, Número 46.
Habermas, Jürguen. 1984. Ciencia y técnica como ideología. Tecnos, Madrid.
Leff, Enrique. 1986. Ecología y capital. Centro de Investigaciones Interdisciplinarias en Ciencias y Humanidades, unam y Siglo XXI Editores, México.
Lyotard, Jean Francois. 1984. La Condición Postmoderna. Cátedra, Barcelona.
Roqueplo, Philippe. 1983. El reparto del saber. Gedisa, Barcelona.
Sagan, Carl. 1997. El mundo y sus demonios. Planeta, Línea Horizonte, México.
Salinas Amescua, B., P. Gochicoa y E. Safa Barraza. 1995. “Educación de adultos y educación popular”, en Educación, cultura y procesos sociales, West Silvia Ma. Teresa (coordinadora). Consejo Mexicano de Investigación Educativa A. C., Coordinación de Humanidades, unam, México.
Sánchez Vázquez, Adolfo. 1996. Izquierda y derecha en política: ¿Y en la moral? cch-sur, unam, México.
Villoro, Luis. 1982. Creer, saber, Conocer. Siglo XXI Editores, México.
|
||
|
Miguel Fernando Pacheco Muñoz
Coordinador de Programas Educativos, Dirección General de Zoológicos de la Ciudad de México.
_______________________________________________________________
como citar este artículo → Pacheco Muñoz, Miguel Fernando. (2003). La divulgación de la ciencia en los tiempos de postmodenidad. Ciencias 71, julio-septiembre, 56-64. [En línea] |
||
| ←Regresar al índice / artículos siguientes→ | ||




