
")

 |
 |
|
| Principales líneas de estudio en la biología evolutiva contemporánea |
|
|
|
Douglas J. Futuyma
conoce más del autor
|
||
| HTML↓ | ||
|
|
||
|
Actualmente vivimos un momento sumamente excitante en la biología evolutiva de los últimos cincuenta o sesenta años, y lo es simplemente porque ha habido un enorme progreso en casi cada aspecto del estudio de la evolución. Este progreso se debe a varios factores: por principio de cuentas se debe a avances tecnológicos tales como el poder secuenciar el adn, poder estudiar la expresión génica o la capacidad de alcanzar la transformación genética pasando genes de un organismo a otro. La aceleración en dicho progreso es el resultado de avances en la tecnología de la información. Hoy día contamos con bases de datos como los bancos de genes y poseemos una enorme capacidad de cómputo que hace posible resolver problemas como las relaciones filogenéticas de grandes grupos de organismos que no hubieran podido ser resueltas en el pasado. También existen avances en la teoría del cambio evolutivo, por ejemplo la teoría de la coalescencia, que es un aspecto enormemente importante y nuevo en genética de poblaciones. Incluso hay cambios en la forma en que podemos analizar los datos evolutivos. Hoy día también poseemos métodos estadísticos sofisticados para mapear los loci de rasgos cuantitativos (lrc o qtl por sus siglas en inglés), lo cual significa que podemos encontrar aquellos loci que contribuyen a la variación en las características poligénicas. Además existen cambios sociológicos que añaden avances a la biología evolutiva, por ejemplo hay una conjunción entre los estudios microevolutivos y los macroevolutivos.
Finalmente, también existen avances en la biología evolutiva simplemente porque se ha incrementado el poder de comunicación gracias al correo electrónico, a las revistas científicas en línea que hacen posible el intercambio entre biólogos de todo el mundo, y esto en sí mismo contribuye al avance de toda la ciencia. Y ya que el progreso ha tocado casi cada aspecto de la biología evolutiva, me concentraré en los aspectos de mayor interés personal.
Los estudios filogenéticos
La producción de filogenias de distintos grupos de organismos nos proporciona hipótesis acerca de la historia evolutiva y también de los procesos que la han generado.
Déjenme ofrecerles dos usos de la filogenia: el primer ejemplo tiene que ver con establecer cuáles especies están más cercanamente emparentadas en un grupo; es decir cuáles de ellas son “especies hermanas”. Debido a que existe un reloj molecular en el cambio en una secuencia de adn, de tal manera que la diferencia en la misma secuencia entre dos especies hermanas se incrementa de manera más o menos constante a través del tiempo desde el momento en que se separaron, es posible calibrar la tasa de evolución de la secuencia, y esto nos permite, por primera vez, fechar la especiación, es decir establecer cuándo ocurrió la divergencia. La figura 1 muestra un ejemplo interesante. Nancy Nowells, quien fuera mi estudiante, investigó la especiación en el género Melanoplus de la familia Acrididae (Ortóptera). Este es un grupo monofilético de una especie de chapulines que habita en las Montañas Rocallosas en altitudes elevadas. Mi alumna se interesó en el patrón e historia de especiación y encontró varios resultados interesantes. Uno relevante es que la divergencia en la secuencia del adn mitocondrial de estas especies es de menos de 4%. Debido a que existen numerosas calibraciones para insectos en la divergencia en la secuencia de adn mitocondrial, su conclusión es que el grupo completo de especies se originó durante el Pleistoceno, esto es durante los últimos 1.8 millones de años. Como puede apreciarse en la figura 1, han ocurrido cuatro o cinco eventos sucesivos de especiación en un periodo relativamente corto, lo que indica claramente que el proceso de especiación ha sido extremadamente rápido, quizás como consecuencia de la fragmentación de poblaciones debido al avance de los glaciares durante el Pleistoceno.
|
||
 figura 1
|
||
| El segundo ejemplo tiene que ver con la especiación en aves. Cuando estudiaba mi doctorado se aceptaba que las especies de aves de Norteamérica cercanamente emparentadas habían especiado durante el periodo Pleistocénico. Sin embargo esta idea ha sido desafiada recientemente por investigadores como Klicka y Zink. Este último estudió distintos pares de especies hermanas de aves en Estados Unidos y analizó el grado de divergencia entre secuencias de adn. En la figura 2 se muestra el número de especies hermanas de aves comparado con la divergencia en la secuencia entre cada par. Uno esperaría que mientras más antigua sea la separación de dos especies hermanas mayor será la cantidad de divergencia entre ellas. Sin embargo, la conclusión de Klicka y Zink, reforzada por los datos presentados en la figura, indica que la mayoría de los pares de especies hermanas de aves que se pensaba que habían evolucionado recientemente en realidad divergieron durante el Plioceno (entre cinco y dos millones años), de manera que probablemente los eventos de especiación no fueron promovidos por el avance de los glaciares. De esta forma, los ejemplos anteriores nos indican que debemos pensar nuevamente la escala de tiempo a la cual la fauna moderna ha evolucionado. | ||
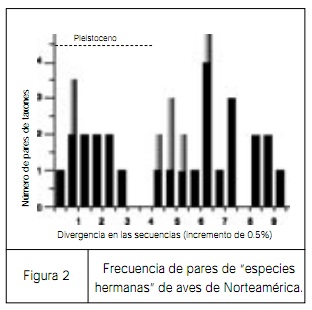 figura 2
|
||
|
Otro aspecto interesante de los estudios filogenéticos es que ahora podemos decir algo acerca de los mecanismos de la especiación. Por ejemplo, los estudios filogenéticos nos permiten describir la polaridad, es decir la dirección de la evolución de una característica. Si una característica tuviera dos estados en un grupo de especies emparentadas, nosotros podemos decir con frecuencia cuál de los estados es el ancestral y cuál es el estado apomórfico o derivado. Hasta hace poco era difícil afirmar esto. Hoy día además es posible decir si un estado derivado ha evolucionado por convergencia o por paralelismo.
Las especies de parásitos o de insectos herbívoros que pueden alimentarse de muchos hospederos se denominan generalistas, mientras que otras especies de parásitos o de insectos herbívoros que se alimentan de sólo una o de unas pocas especies de hospederos se denominan especialistas. Tradicionalmente se ha postulado que la especialización es un “callejón sin salida” evolutivo. Es decir que las especies especialistas no podrían evolucionar para convertirse en cualquier otra cosa y quedan “estancadas” en un nicho ecológico hasta que finalmente se extinguen. En resumen, la noción clásica es que las especies especialistas han derivado o han evolucionado a partir de especies generalistas, y que las especialistas no pueden originar especies generalistas.
Brian Farrell y su estudiante Kelley son de los primeros que han intentado abordar esta pregunta analizando un grupo de escarabajos del género Dendroctonus. Estos escarabajos atacan principalmente especies de coníferas; algunas especies están muy especializadas mientras que otras pueden alimentarse de distintas especies de coníferas. En la figura 3 es posible apreciar la filogenia del género Dendroctonus. Las especies generalistas de Dendroctonus están ilustradas en blanco y las especialistas en negro. Puede apreciarse claramente que las especialistas están siempre al final de las ramas del árbol filogenético, lo cual apoya la hipótesis tradicional de que a partir de las especies generalistas evolucionan las especialistas. Lo contrario no se aprecia en la filogenia del género Dendroctonus, y ésta es la primera vez que hemos sido capaces de poner a prueba la hipótesis tradicional sobre la evolución de los especialistas y generalistas. Esta pregunta es relevante porque está en la frontera de la ecología y la evolución. Sin embargo, después del análisis de Farrell, otros estudios como el de Nosil, en otros grupos de insectos han demostrado que la hipótesis tradicional no siempre es verdadera. Existen algunos grupos de especialistas que pueden dar origen a especies generalistas y algunas veces el fenómeno ocurre en ambas direcciones. De manera que, hasta el momento, no podemos concluir acerca de la evolución del generalismo y el “especialismo” ecológico, pero es probable que el estudio de otros grupos de parásitos haga posible llegar a una conclusión más sólida.
|
||
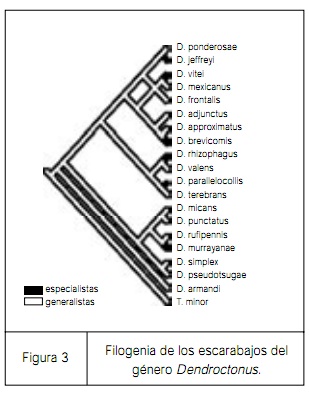 figura 3
|
||
|
La teoría de la coalescencia
El segundo tema importante que me gustaría mencionar es el estudio de lo que llamamos genealogía de genes, es decir filogenias de diferentes secuencias de adn de un locus entre especies o dentro de un grupo de especies cercanamente emparentadas. De manera que podamos trazar un ancestro común entre dos o tres especies, y podamos tomar tres secuencias variables de adn y, por los mismos métodos filogenéticos, llegar a la conclusión de cuáles secuencias de adn dentro de un grupo de especies son las más cercanamente relacionadas. Este hecho, en conjunto con la teoría matemática, es la base de la teoría de la coalescencia. La teoría de la coalescencia ve a la variación genética dentro de una población con una historia, o sea que los diferentes alelos o secuencias de adn dentro de una población tienen una historia, y que en última instancia todas las diferentes secuencias debieron de haberse originado de una secuencia original en el pasado. La mayoría de las secuencias pudieron haberse eliminado por deriva génica o quizás por selección natural, de manera que las secuencias existentes vienen de una secuencia ancestral y podríamos preguntarnos: ¿cuál es la secuencia ancestral?, ¿es reciente o es muy antigua? Y si pudiéramos tener esa estimación de la edad, entonces podríamos hacer inferencias acerca de la selección sobre esa secuencia, ya sea selección balanceadora o selección direccional. Algunas veces incluso es posible estimar el tamaño histórico de la población simplemente por el hecho de que una población que ha sido de tamaño muy grande, preservará las distintas secuencias por más tiempo, debido a que éstas no se extinguen rápidamente por deriva génica. También es posible, algunas veces, realizar una estimación de si la población ha crecido en tamaño o ha permanecido del mismo tamaño a través del tiempo.
Otro aspecto a estudiar de las genealogías de genes es la distribución geográfica de las diferentes variantes de un gen, que puede ofrecer una estimación de la historia de los cambios en la distribución de las poblaciones que constituyen la especie. Este es el campo de la filogeografía, un campo muy activo en el pasado reciente y que promete aportar una gran cantidad de información a la biología evolutiva. Veamos dos ejemplos.
Los genes de autoincompatibilidad en plantas evitan que un genotipo pueda producir descendencia por autopolinización. El árbol ilustrado en la figura 4 corresponde a distintos haplotipos, es decir, diferentes secuencias de un locus que produce autoincompatibilidad en las plantas de la familia Solanaceae. Lo que se observa en la figura 4 son diferentes formas o alelos del mismo gen, y puede apreciarse que en Nicotiana existen diferentes alelos de autoincompatibilidad. Hay cuatro diferentes alelos en este locus, y por lo tanto se dice que es un locus polimórfico, lo mismo puede decirse para el género Petunia. Pero lo relevante de este ejemplo es que los alelos del género Nicotiana (nic) están más cercanamente relacionados, no con otros del mismo género, sino con otros alelos que se encuentran en Solanum (sol), Petunia (pet) o Lycopersicon. Lo que significa que se trata de un polimorfismo muy antiguo, del cual ha habido linajes de genes por mucho tiempo y que todos, o varios de estos linajes de genes, fueron heredados por Nicotiana, Petunia o Solanum. En resumen, quiere decir que este polimorfismo ha existido desde el ancestro común de todos estos géneros de Solanaceae. Otra pregunta que podríamos hacernos es: ¿Qué tan antiguo es o por cuánto tiempo ha persistido este polimorfismo? La respuesta no la sabemos con certeza, pero el origen podría remontarse al Mioceno, de 30 a 40 millones de años. Y esto significa que probablemente ha estado bajo selección balanceadora, es decir una selección que ha mantenido el polimorfismo en la población. Hoy, por ejemplo, sabemos que opera la selección dependiente de la frecuencia sobre los alelos de autoincompatibilidad en plantas; sin embargo, lo que no sabíamos es lo poderosa que puede ser la selección para mantener un polimorfismo por 30 millones de años o más. Éste es un hallazgo sorprendente.
|
||
 figura 4
|
||
| El segundo ejemplo acerca del tipo de inferencias que podemos obtener de una filogenia de genes está relacionado con nosotros mismos, los seres humanos. La figura 5 muestra un árbol filogenético derivado a partir de la variación en secuencias de adn mitocondrial en humanos y en chimpancés. En el árbol tenemos una secuencia para chimpancés y varias para humanos de diferentes partes del mundo, de donde provienen estas secuencias. Lo primero que observamos es que todas las secuencias antiguas están presentes en África, y la longitud de las ramas indica la cantidad de diferencias mutacionales que existen entre las secuencias. Los resultados también indican que las secuencias son muy antiguas, o sea que han permanecido por mucho tiempo, el suficiente para acumular diferentes mutaciones, y por lo tanto indican claramente que las secuencias más antiguas existieron en África y después se extendieron al resto del mundo. De manera que tenemos evidencia incontrovertible de que África fue el hogar original de Homo sapiens, lo que por supuesto ya conocíamos por el registro fósil; en segundo lugar sabemos que los humanos han habitado África por mucho más tiempo que ninguna otra parte del mundo y que en ese continente los humanos han tenido una población más grande que en el resto del mundo. También podemos apreciar, gracias al adn mitocondrial, que las secuencias de otras partes del mundo son casi idénticas. Han ocurrido muy pocas diferencias mutacionales de un lugar a otro, como se aprecia por lo corto de las ramas. De manera que todas las secuencias mitocondriales debieron haber descendido de una secuencia ancestral común en el pasado reciente y posteriormente de grupos poblacionales pequeños que colonizaron el resto del mundo. Este aspecto es uno de los más interesantes que se han descubierto acerca de la historia evolutiva de nuestra especie. Esta aproximación también puede ser empleada para el estudio de otras especies. | ||
 figura 5
|
||
| Otro aspecto derivado de la teoría de la coalescencia y que se relaciona con el tamaño poblacional ancestral y la retención de variantes o de secuencias de genes, se puede apreciar en la figura 6a, que muestra una genealogía de genes relacionados entre sí y que difieren sólo en cambios mutacionales, pero todos derivan de un ancestro común. Cuando la población sea muy grande, todos los linajes de genes serán retenidos en la población por largo tiempo y tendrán muchos genes que estarán muy cercanamente relacionados, por lo que sólo existirán pequeñas diferencias entre ellos. Mientras más diferencias existan entre dos secuencias, más lejano en el pasado estará el ancestro común. Pero, debido a que la población es de tamaño grande, estos linajes de genes se mantendrán por mucho tiempo. De manera que si tomamos pares de secuencias de adn para este locus y calculamos el porcentaje de pares que tienen cero, una, dos, tres o más diferencias, esperaríamos una distribución de frecuencias como la que se ilustra en la figura 6a. Sin embargo, si la población ha sido pequeña y ha pasado por “cuellos de botella” poblacionales, es probable que los genes que existen hoy día vengan de un ancestro común reciente. Si la población está en expansión acumulará diferencias mutacionales, pero debido a que no ha habido suficiente tiempo para crear un árbol completo, la distribución de las frecuencias de las diferencias entre pares de genes será totalmente distinta a aquella derivada de poblaciones de tamaño grande (figura 6b). | ||
 figura 6
|
||
|
Por supuesto, si aplicamos esto a las poblaciones de humanos en Asia y en Europa, ambas aparecen como si hubieran sido originadas por poblaciones pequeñas.
Dicha teoría se puede aplicar a la evolución de nuevas especies. Uno de los postulados acerca de cómo surgen las especies fue hecho por Ernst Mayr en 1954, denominado teoría de la especiación por efecto del fundador. Mayr postuló que una nueva especie frecuentemente tiene su origen en poblaciones pequeñas, fundadas por pocos individuos. Si esto es verdad, será posible apreciar en las especies así originadas la “firma” de un cuello de botella en la variación en las secuencias de adn. Precisamente esto fue lo que estudiamos en el escarabajo Ophraella. En la filogenia de este escarabajo, figura 7, tenemos a las especies Ophraella communa, ampliamente distribuida desde el sur de Canadá hasta Guatemala, y Ophraella bilineata, distribuida únicamente en el borde de la distribución de la primera. Ambas están muy cercanamente emparentadas y supusimos que quizás Ophraella bilineata se originó en el borde de la distribución, justo como Ernst Mayr postuló para el origen de muchas especies nuevas. Nos preguntamos si la especie Ophraella bilineata mostraría una reducción en la variación genética y una distribución única de las frecuencias de las secuencias. Los resultados indican claramente que la población periférica no muestra evidencia de haberse originado por efecto de un fundador, y por lo tanto no apoyan la hipótesis de Ernst Mayr sobre el origen de nuevas especies a partir de un pequeño número de individuos. De manera que la teoría de la coalescencia puede usarse para explorar algunos aspectos de la especiación. Si una población ha sido fundada por unos cuantos individuos, uno esperaría que tuviera un número pequeño de secuencias diferentes, secuencias de diferentes haplotipos que probablemente estarían cercanamente relacionados uno con otro y que difirieran sólo en unas cuantas mutaciones. Aquí es donde entra la filogeografía, es decir la distribución geográfica de una secuencia de genes en relación a la filogenia de los genes.
|
||
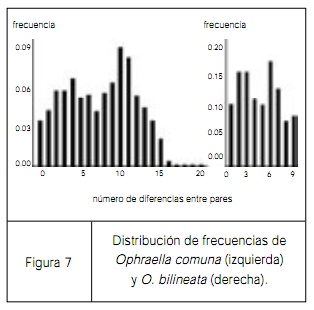 figura 7
|
||
|
La filogeografía
El siguiente ejemplo es acerca de un ave pequeña, Oporornis tolmiei, cuya distribución geográfica se presenta en la figura 8. Esta ave tiene una población reproductiva en un área enorme del oeste de Estados Unidos y en Canadá. En la figura también se muestra otra área en el este de Coahuila y oeste de Nuevo León; un área mucho más pequeña de una población disyunta con respecto a la costa oeste de Norteamérica. Se analizaron tanto la población mexicana como la de Estados Unidos, usando secuencias de adn mitocondrial, y se construyó el árbol de genes y de las localidades de las cuales se tomaron las muestras. Los resultados indican que hay un haplotipo que es muy común en los Estados Unidos y está presente en casi todas las poblaciones al igual que en México. Mientras que este haplotipo es el más común y es casi el único haplotipo en Estados Unidos, aunque tiene algunas variantes que difieren por mutaciones únicas o sencillas, las muestras tomadas de la población reproductiva en México son mucho más variables en las secuencias de adn mitocondrial; aquí existen múltiples haplotipos que son igualmente frecuentes y que tienen hasta cuatro diferencias mutacionales.
|
||
 figura 8
|
||
|
Esto nos indica que este grupo de haplotipos es mucho más antiguo y ha persistido por mucho más tiempo en México que en Estados Unidos. La distribución de frecuencias de las diferencias en las secuencias de la población de los Estados Unidos es unimodal, lo cual sugiere que esta población creció recientemente a partir de un número pequeño de individuos. Por el contrario, la distribución de frecuencias de la población mexicana tiene una forma que sugiere que la población original ha sido grande. ¿Qué quiere decir ésto? Esto sugiere que la población de esta ave en México es la población ancestral y que probablemente en la actualidad es una población relicto en esta área geográfica de México. Esta evidencia indica que la historia de las especies y sus distribución geográfica pueden ser inferidas a partir de árboles de genes.
La especiación es muy difícil de estudiar debido a que usualmente es un proceso sumamente lento para ser observado, pero también es temporalmente muy corto como para quedar grabado en el registro fósil. Por ello, casi cada aspecto de la especiación es controvertido. Pero hoy existe la esperanza de poder responder a las viejas preguntas acerca de la especiación. Por ejemplo, hoy poseemos nuevas herramientas para analizar las diferencias genéticas entre especies, de manera que podemos abordar el análisis del aislamiento reproductivo entre especies, su aislamiento conductual, mecánico o fisiológico y saber si producen híbridos y éstos son estériles. Hoy también podemos preguntarnos si el aislamiento reproductivo es producido por una diferencia de uno, dos o tres genes, o si ésta requiere que haya diferencias en muchos más genes. Intentamos conocer cuánto tiempo toma el proceso de especiación. Tenemos indicios, como los de los estudios de los chapulines y de las aves, de que el proceso de especiación a veces puede ser muy rápido.
También podemos preguntarnos si la especiación simpátrica es posible, es decir aquella que ocurre cuando no hay barreras en el flujo genético entre las poblaciones que ocupan una misma área. También he mostrado que podemos preguntarnos si la especiación por el efecto fundador es probable o no. Podemos tener alguna guía acerca de qué tan frecuentemente surgen las especies por el efecto fundador, justo como lo sugirió Ernst Mayr, pero una pregunta que podríamos abordar y que casi nadie ha contestado sino hasta muy recientemente es la siguiente: ¿Es la especiación producida por selección natural? La mayoría de los evolucionistas ha supuesto siempre que el aislamiento reproductivo es producido por la selección natural. Una idea popular entre los evolucionistas, sugerida por Ernst Mayr y Theodosius Dobzhansky, es que las poblaciones geográficas de una especie se adaptan a diferentes condiciones ecológicas y que los cambios genéticos tendrán efectos pleiotrópicos, es decir efectos colaterales que causan alteraciones en el comportamiento, desarrollo o fisiología, y que las hacen incompatibles con otras poblaciones, por lo cual no pueden intercambiar genes, ni se cruzan o son incapaces de producir descendencia viable y fértil. En resumen, la idea es que la especiación es un producto secundario de la selección natural ecológica; la selección, al adaptar localmente a las poblaciones, favorece indirectamente el aislamiento reproductivo.
Pero si uno revisa los libros de texto, incluida la primera edición de mi libro Biología evolutiva, y busca ejemplos acerca del papel de la selección natural en el origen de nuevas especies o del aislamiento reproductivo, difícilmente encontrará uno debido a que nadie se había fijado en esa pregunta sino hasta hace muy poco. Aunque todo mundo asume que la selección natural produce especiación, el hecho es que no lo sabemos con certeza. ¿Es posible que dos poblaciones que están aisladas sufran cambios por deriva génica, es decir cambios puramente aleatorios, y se vuelvan los suficientemente distintas genéticamente, de manera que sean incompatibles una con otra? Esto es totalmente plausible. De hecho sabemos muy poco acerca de si la selección natural o la deriva génica son responsables de la especiación. Hasta hace poco sólo un pequeño número de investigadores había entendido lo importante que es esta pregunta: ¿Qué es lo que causa la evolución de nuevas especies? Es decir, ¿qué puede ser más importante que eso? Uno de mis estudiantes, Daniel Funk, intentó contestar la siguiente pregunta: ¿La adaptación a diferentes condiciones ecológicas acelera la evolución del aislamiento reproductivo? Para ello, estudió insectos herbívoros, especializados en diferentes especies de plantas, y se preguntó si en poblaciones de una especie adaptadas a diferentes especies de plantas hospederas, la adaptación divergente acelera el origen del aislamiento reproductivo entre aquellas poblaciones geográficas. Supongamos que tenemos dos poblaciones separadas y que ambas se alimentan de la misma planta A (la población 1 y la población 2). Es probable que exista cierta divergencia genética entre ambas poblaciones debido a distintas fuerzas evolutivas, algo de deriva génica, quizás diferencias producidas por el clima o por diferentes depredadores, es decir, divergencia genética causada por otras causas distintas al aislamiento reproductivo. Pero si tenemos dos poblaciones que ocupan diferentes plantas hospederas, entonces habrá las mismas fuerzas evolutivas misceláneas que dan origen a la divergencia genética, además de las diferencias que surgen por su adaptación a diferentes plantas hospederas. Y si la hipótesis de Mayr de que el aislamiento reproductivo evoluciona como un efecto colateral de la divergencia genética debido a las circunstancias ecológicas es cierta, entonces en las diferentes poblaciones asociadas a un hospedero distinto evolucionará más rápido el aislamiento reproductivo que en las poblaciones que ocupan el mismo hospedero. Para poner a prueba esta hipótesis, Funk eligió una especie de escarabajo crisomélido del género Neochlamisus, en particular la especie Neochlamisus bebbianae. Este escarabajo se alimenta de tres especies de plantas; en gran parte del este de Norteamérica así como en Ontario, Canadá, se alimenta del maple rojo (Acer rubrum). Hay otras poblaciones que por el contrario se alimentan de sauces (género Salix) y las de Oklahoma se alimentan del abedul (género Betula). Funk tomó poblaciones que se alimentaban del maple rojo en Nueva York y en Georgia como el par de poblaciones que se alimentan del mismo hospedero; también tomó poblaciones que se alimentaban de Salix y también de Betula. Lo que hizo entonces en el laboratorio fue poner parejas en cajas de Petri: macho y hembra provenientes del mismo o diferente hospedero y observó su comportamiento. Registró el porcentaje de parejas que copularon y produjeron huevos; por otra parte, también demostró que estas poblaciones de insectos están adaptadas a diferentes plantas hospederas, de manera que las poblaciones que se alimentan de Salix tienen una mejor adaptación a este hospedero que los escarabajos que se alimentan de Acer. Una medida del aislamiento sexual es la proporción de parejas que no se aparean. El primer resultado fue que las poblaciones de Georgia que se alimentan de maple no mostraron aislamiento sexual con las poblaciones de Nueva York, que también se alimentan de maple, es decir que no existe barrera reproductiva entre ellas. Pero cuando tomaba poblaciones de Georgia o de Nueva York que se alimentan de maple y las ponía junto al sexo opuesto de las poblaciones de Ontario que se alimentan de Salix, lo que encontró fue un alto grado de aislamiento reproductivo. Lo mismo ocurrió cuando comparó a los escarabajos que se alimentan de Acer con los que se alimentan de Betula: cuando puso un macho proveniente de Betula con una hembra de Acer o viceversa encontró un alto grado de aislamiento sexual. De manera que, tal como se predijo, el aislamiento sexual es más alto entre poblaciones que están adaptadas a diferentes plantas hospederas. Funk también analizó el adn mitocondrial debido a que las diferentes poblaciones de diferentes hospederas pudieran ser más viejas, y de esta manera la divergencia se explicaría por el tiempo durante el cual se han acumulado las diferencias genéticas por deriva génica. Pero lo que encontró fue que las dos poblaciones que se alimentan de Acer eran más viejas; es decir que tuvieron un antepasado común más antiguo que el de Salix de Canadá, y por lo tanto hay menos tiempo de divergencia entre la población de escarabajos que se alimenta de Salix que de la población que se alimenta de Acer en Nueva York. Los resultados de este estudio apoyan la hipótesis de Mayr y Dobzhansky sobre la especiación o el aislamiento reproductivo como un producto secundario de la adaptación ecológica a diferentes circunstancias; en este caso plantas hospederas.
Evolución de las interacciones ecológicas
¿Cómo interactúan las especies una con otra, ya sea depredador y presa, parásito y hospedero o mutualistas y competidores? ¿Cómo evolucionan en conjunto? Y es aún más compleja la pregunta, ¿cómo es que la estructura de una comunidad ecológica depende de la evolución?
Veamos brevemente algo acerca de la evolución del uso de recursos, ya que éste es un tópico muy importante para la interacción de especies y subyace en gran parte de la teoría de la ecología de las comunidades. Cuando estudiaba mi doctorado, Robert MacArthur y su mentora Evelyn Hutchinson eran los ecólogos más importantes y creían que era posible producir una teoría general de la estructura de las comunidades, basada parcialmente en la teoría de la evolución. La idea era que, por ejemplo, las especies que compiten evolucionarían para segregarse en lo que llamamos nichos ecológicos, de tal forma que habría una explotación consistente o una división de los recursos entre un cierto número de especies que minimizaría la cantidad de competencia entre ellas. La evidencia más clara de apoyo a esta idea proviene del trabajo de Jim Brown, quien mostró que en dos desiertos de Norteamérica las especies de roedores de diferentes tamaños, que se alimentan de semillas, en gran medida están segregadas en diferentes clases de recursos en relación al tamaño del roedor. La comparación entre las comunidades mostró que había un paralelismo, es decir, una correspondencia de especies en ambos desiertos, y esto fue tomado como el ejemplo de que la estructura de la comunidad puede predecirse y que puede evolucionar a una clase consistente, casi óptima, como una función de la evolución de estas especies con otras.
Pero una de las grandes preguntas, que considero aún permanece en ecología evolutiva, es si este cuadro optimista de la predecibilidad de las comunidades es verdadero o no. Y la respuesta parece ser que algunas veces lo es y otras no. Veamos lo que considero como el ejemplo más espectacular en apoyo a la teoría de Robert MacArthur: las lagartijas del género Anolis en las Antillas. En las grandes islas, Cuba, Puerto Rico, Haití y Dominicana, y en Jamaica, existen varias especies de lagartijas del género Anolis que se segregan en distintos hábitats, de manera que unas viven en las copas de los árboles, otras viven en el tronco o corteza, algunas en pequeñas ramas e incluso las hay que viven en el suelo o en la base de los árboles. Pero el hecho más notable es que en la filogenia de las especies de Anolis en cada una de las Grandes Antillas encontramos similitudes impresionantes: en primer lugar, el hecho ecológico es que las Anolis están segregadas en diferentes hábitats, pero la filogenia indica que en cada isla el grupo de Anolis es monofilético. De manera que esto implica que Anolis ha irradiado, en respuesta al mismo conjunto de circunstancias en el uso de recursos; este es un caso de radiación adaptativa, y es extraordinario porque muestra la predecibilidad de la evolución entre especies que compiten y se segregan en distintos nichos de manera consistente. Sin embargo este fenómeno no siempre ocurre. Por ejemplo, los escarabajos de la tribu Tetraopini (familia Cerambicidae, orden Coleoptera), cuyo origen se remonta al Oligoceno (35 millones de años), se alimentan sólo de plantas de la familia Apocynaceae, y esta restricción histórica de un grupo de insectos a un grupo particular de plantas indica que estos insectos no han sido capaces de adaptarse rápidamente a diferentes clases de hospederos. A diferencia de las especies de lagartijas Anolis, que pueden reaccionar a la presencia de competidores evolucionando un cambio en la forma en la cual usan los recursos disponibles, parece que muchos grupos de insectos no tuvieran esa capacidad para evolucionar hacia un cambio de su planta hospedera particular, de forma que pudieran expandirse a distintas especies de plantas en una comunidad. El resultado de esto es que en muchos grupos de insectos existe un impacto de la historia evolutiva a largo plazo en la estructura contemporánea de la comunidad de plantas y de insectos. Esto da como resultado que ciertas especies de plantas tengan menos clases de insectos alimentándose de ellas, mientras que otras especies de plantas tendrán muchos insectos. Y esto es, en parte, una cuestión que tiene que ver con la cantidad de tiempo evolutivo en el cual ha habido oportunidad para los insectos de adaptarse a las plantas cuando son nuevas para ellos. Por ejemplo, la familia de plantas Asteraceae tiene muchas tribus distintas; algunas de estas tribus llegaron a Norteamérica hace sólo unos cuantos millones de años y tienen una muy pequeña fauna de insectos alimentándose de ellas; sin embargo, otras tribus como Astereae y Heliantheae son mucho más viejas en Norteamérica y tienen un vasto número de especies asociadas. De manera que la estructura contemporánea de la cadena alimentaria depende en gran medida de qué tan antigua sea la historia evolutiva en el caso de las interacciones de planta e insecto, mientras que en el caso de las lagartijas del género Anolis ha ocurrido un acomodamiento evolutivo de una especie a otra.
Epílogo
En los párrafos precedentes he tratado de señalar algunas de las preguntas que se analizan en ciertas áreas de la biología evolutiva en las cuales estamos atestiguando un grado de progreso y una profundidad en el entendimiento que no habíamos alcanzado antes. Si miro a las ediciones anteriores de mi libro de texto, Biología evolutiva, y comparo lo que dice con lo que sabemos hoy, me digo, ¡por Dios!, ¿yo escribí esto? Es tan diferente de lo que sabemos hoy que es casi como otro siglo. Y este progreso tiene implicaciones importantes fuera del ámbito académico del estudio de la evolución debido a que los biólogos evolucionistas, como muchos otros biólogos, están entendiendo que tienen cosas importantes que decir que son de valor social para la comunidad como un todo. Tenemos muchas cosas que decir acerca de la evolución de los parásitos, incluidos los microbios y los patógenos; tenemos muchas cosas importantes que decir acerca del papel de los genes en el desarrollo y los efectos pleiotrópicos de los genes en la especiación. El análisis de los efectos genéticos sobre el fenotipo es importante si uno empieza a pensar sobre los organismos transgénicos o genéticamente modificados por humanos. Existen muchas formas en las cuales la biología evolutiva es aplicable a cuestiones sociales de nuestra era, desde la salud humana hasta la conservación de las especies que todos apreciamos. Por lo tanto, quiero reiterar que éste es uno de los tiempos más excitantes para ser un biólogo evolutivo y me congratulo de que muchos de quienes estén aquí escuchado lo sean.
|
||
|
Traducción
Juan Núñez Farfán
Conferencia presentada en la Facultad de Ciencias el 28 de noviembre de 2002.
Transcripción: Mónica Benítez.
|
||
|
Referencias bibliográficas
Funk D. J. “Isolating a role for natural selection in in speciation: host adaptation and sexual isolation in Neochalamisus Bibbianae haf Beattlis”, en Evolution, núm. 52, pp. 1744-1759.
Futuyma, D. J. 1979. Evolutionary biology. Sunderland, Massachussets.
Ingman, M., H. Kaessmann, S. Paabo y U. Gyllensten. 2000. "Mitochondrial genome variation and the origin of modern humans", en Nature, núm. 408, pp. 708-713.
Ioerger, T. R., A. G. Clark y T. H. Kao. 1990. "Polymorphism at the self-incompatibility locus in Solanaceae predates speciation", en Proceedings of the National Academy of Sciences, Estados Unidos, núm. 87, pp. 9732-9735.
Kelley, S. T. y B. D. Farrell. 1998. "Is specialization a dead end? The phylogeny of host use in Dendroctonus bark beetles (Scolytidae)", en Evolution, núm. 52, pp. 1731-1743.
Klicka, J., and R. M. Zink. 1997. "The importance of recent ice ages in speciation: a failed paradigm", en Science, núm. 277, pp. 1666-1669.
Knowles, L. L., D. J. Futuyma, W. F. Eanes y B. Rannala. 1999. “Insight into speciation from historical demography in the phytophagous beetle genus Ophraella”, en Evolution, núm. 53, pp. 1846-1856.
Knowles, L. L. 2000. “Tests of Pleistocene speciation in montane grasshoppers (genus Melanoplus) from the sky islands of western North America”, en Evolution, núm. 54, pp. 1337-1348.
Mila et al. 2000. “Genetic evidence for ten effect of a postglacial population expansion on the phylogeography of a Northamerican Sungbird”, en Proceedings of the Royal Society of London, núm. B 267, pp. 1033-1040.
Nosil, P. 2002. "Transition rates between specialization and generalization in phytophagous insects" en Evolution, núm. 56, pp. 1701-1706.
Figuras
1. Cortesía de L. L. Knowles; 2. Klicka y Zink, 1999; 3. Kelley y Farell, 1998; 4. Ioerger et. al. 1990; 5. Ingman et. al. 2000; 6. Cortesía de Ingman et. al. 2000; 7. Cortesía de L. L. Knowles, 8. Mila et. al. 2000.
|
||
|
Douglas J. Futuyma
Departamento de Ecología y Biología Evolutiva, Universidad de Michigan.
_______________________________________________________________
como citar este artículo → Futuyma, Douglas J. (2003). Principales líneas de estudio en la biología evolutiva contemporánea. Ciencias 71, julio-septiembre, 16-28. [En línea] |
||
| ←Regresar al índice / artículo siguiente→ | ||
revista de cultura científica FACULTAD DE CIENCIAS, UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
Busca ampliar la cultura científica de la población, difundir información y hacer de la ciencia
un instrumento para el análisis de la realidad, con diversos puntos de vista desde la ciencia.
You are here: Inicio  Búsqueda Número revistas revista ciencias 71 Principales líneas de estudio en la biología evolutiva contemporánea
Búsqueda Número revistas revista ciencias 71 Principales líneas de estudio en la biología evolutiva contemporánea
|
Universidad Nacional Autónoma de México
Facultad de Ciencias Departamento de Física, cubículos 320 y 321.
Ciudad Universitaria. México, D.F., C.P. 04510. Télefono y Fax: +52 (01 55) 56 22 4935, 56 22 5316 Trabajo realizado con el apoyo de: Programa UNAM-DGAPA-PAPIME número PE103509 y UNAM-DGAPA-PAPIME número PE106212 |
ISSN:0187-6376 Responsable del sitio Laura González Guerrero This email address is being protected from spambots. You need JavaScript enabled to view it. Asesor técnico: e-marketingservices.com |
This work is licensed under a Creative Commons Attribution-Noncommercial-Share Alike 3.0 United States License |
![]()